Этапы развития нервной системы. Развитие нервной системы. Различают такие отростки твердой мозговой оболочки
Нервная система имеет эктодермальное происхождение, т. е. развивается из внешнего зачаточного листка толщиной в одноклеточный слой вследствие образования и деления медуллярной трубки. В эволюции нервной системы схематично можно выделить такие этапы.
1. Сетевидная, диффузная, или асинаптическая, нервная система. Возникает она у пресноводной гидры, имеет форму сетки, которая образуется соединением отростчатых клеток и равномерно распределяется по всему телу, сгущаясь вокруг ротовых придатков. Клетки, которые входят в состав этой сетки, существенно отличаются от нервных клеток высших животных: они маленькие по размеру, не имеют характерного для нервной клетки ядра и хроматофильной субстанции. Эта нервная система проводит возбуждения диффузно, по всем направлениям, обеспечивая глобальные рефлекторные реакции. На дальнейших этапах развития многоклеточных животных она теряет значение единой формы нервной системы, но в организме человека сохраняется в виде мейснеровского и ауэрбаховского сплетений пищеварительного тракта.
2. Ганглиозная нервная система (в червеобразных) синаптическая, проводит возбуждение в одном направлении и обеспечивает дифференцированные приспособительные реакции. Этому отвечает высшая степень эволюции нервной системы: развиваются специальные органы движения и рецепторные органы, в сетке возникают группы нервных клеток, в телах которых содержится хроматофильная субстанция. Она имеет свойство распадаться во время возбуждения клеток и восстанавливаться в состоянии покоя. Клетки с хроматофильной субстанцией располагаются группами или узлами ганглиями, поэтому получили название ганглиозных. Итак, на втором этапе развития нервная система из сетевидной превратилась в ганглиозно-сетевидную. У человека этот тип строения нервной системы сохранился в виде паравертебральных стволов и периферических узлов (ганглиев), которые имеют вегетативные функции.
3. Трубчатая нервная система (в позвоночных) отличается от нервной системы червеобразных тем, что в позвоночных возникли скелетные моторные аппараты с поперечно-полосатыми мышцами. Это обусловило развитие центральной нервной системы, отдельные части и структуры которой формируются в процессе эволюции постепенно и в определенной последовательности. Сначала из каудальной, недифференцированной части медуллярной трубки образуется сегментарный аппарат спинного мозга, а из передней части мозговой трубки вследствие кефализации (от греч. kephale - голова) формируются основные отделы головного мозга. В онтогенезе человека они последовательно развиваются по известной схеме: сначала формируются три первичных мозговых пузыря: передний (prosencephalon), средний (mesencephalon) и ромбовидный, или задний (rhombencephalon). В дальнейшем из переднего мозгового пузыря образуются конечный (telencephalon) и промежуточный (diencephalon) пузыри. Ромбовидный мозговой пузырь также фрагментируется на два: задний (metencephalon) и продолговатый (myelencephalon). Таким образом, стадия трех пузырей сменяется стадией образования пяти пузырей, из которых формируются разные отделы центральной нервной системы: из telencephalon большие полушария мозга, diencephalon промежуточный мозг, mesencephalon - средний мозг, metencephalon - мост мозга и мозжечок, myelencephalon - продолговатый мозг.
Эволюция нервной системы позвоночных обусловила развитие новой системы, способной образовывать временные соединения функционирующих элементов, которые обеспечиваются расчленением центральных нервных аппаратов на отдельные функциональные единицы нейроны. Следовательно, с возникновением скелетной моторики в позвоночных развилась нейронная цереброспинальная нервная система, которой подчинены более древние образования, что сохранились. Дальнейшее развитие центральной нервной системы обусловило возникновение особых функциональных взаимосвязей между головным и спинным мозгом, которые построены по принципу субординации, или соподчинения. Суть принципа субординации состоит в том, что эволюционно новые нервные образования не только регулируют функции более древних, низших нервных структур, а и соподчиняют их себе путем торможения или возбуждения. Причем субординация существует не только между новыми и древними функциями, между головным и спинным мозгом, но и наблюдается между корой и подкоркой, между подкоркой и стволовой частью мозга и в определенной степени даже между шейным и поясничным утолщениями спинного мозга. С появлением новых функций нервной системы древние не исчезают. При выпадении новых функций появляются древние формы реакции, обусловленные функционированием более древних структур. Примером может служить появление субкортикальных или стопных патологических рефлексов при поражении коры большого мозга.
Таким образом, в процессе эволюции нервной системы можно выделить несколько основных этапов, которые являются основными в ее морфологическом и функциональном развитии. Из морфологических этапов следует назвать централизацию нервной системы, кефализацию, кортикализацию в хордовых, появление симметричных полушарий - у высших позвоночных. В функциональном отношении эти процессы связаны с принципом субординации и возрастающей специализацией центров и корковых структур. Функциональной эволюции соответствует эволюция морфологическая. При этом филогенетически более молодые структуры мозга являются более ранимыми и в меньшей степени обладают способностью к восстановлению.
Нервная система имеет нейронный тип строения, т. е. состоит из нервных клеток - нейронов, которые развиваются из нейробластов.
Нейрон является основной морфологической, генетической и функциональной единицей нервной системы. Он имеет тело (перикарион) и большое количество отростков, среди которых различают аксон и дендриты. Аксон, или нейрит, - это длинный отросток, который проводит нервный импульс в направлении от тела клетки и заканчивается терминальным разветвлением. Он всегда в клетке лишь один. Дендриты - это большое количество коротких древообразных разветвленных отростков. Они передают нервные импульсы по направлению к телу клетки. Тело нейрона состоит из цитоплазмы и ядра с одним или несколькими ядрышками. Специальными компонентами нервных клеток являются хроматофильная субстанция и нейрофибриллы. Хроматофильная субстанция имеет вид разных по размерам комочков и зерен, содержится в теле и дендритах нейронов и никогда не выявляется в аксонах и начальных сегментах последних. Она является показателем функционального состояния нейрона: исчезает в случае истощения нервной клетки и восстанавливается в период покоя. Нейрофибриллы имеют вид тонких нитей, которые размещаются в теле клетки и ее отростках. Цитоплазма нервной клетки содержит также пластинчатый комплекс (сетчатый аппарат Гольджи), митохондрии и другие органоиды. Сосредоточение тел нервных клеток формируют нервные центры, или так называемое серое вещество.
Нервные волокна - это отростки нейронов. В границах центральной нервной системы они образуют проводящие пути - белое вещество мозга. Нервные волокна состоят из осевого цилиндра, который является отростком нейрона, и оболочки, образованной клетками олигодендроглии (нейролемоцитами, шванновскими клетками). В зависимости от строения оболочки, нервные волокна делятся на миелиновые и безмиелиновые. Миелиновые нервные волокна входят в состав головного и спинного мозга, а также периферических нервов. Они состоят из осевого цилиндра, миелиновой оболочки, нейролемы (шванновской оболочки) и базальной мембраны. Мембрана аксона служит для проведения электрического импульса и в участке аксональных окончании выделяет медиатор, а мембрана дендритов - реагирует на медиатор. Кроме того, она обеспечивает распознавание других клеток в процессе эмбрионального развития. Поэтому каждая клетка отыскивает определенное ей место в сети нейронов. Миелиновые оболочки нервных волокон не сплошные, а прерываются промежутками сужений - узлами (узловые перехваты Ранвье). Ионы могут проникать в аксон только в области перехватов Ранвье и в участке начального сегмента. Безмиелиновые нервные волокна типичны для автономной (вегетативной) нервной системы. Они имеют простое строение: состоят из осевого цилиндра, нейролеммы и базальной мембраны. Скорость передачи нервного импульса миелиновыми нервными волокнами значительно выше (до 40-60 м/с), чем немиелиновыми (1-2 м/с).
Основными функциями нейрона являются восприятие и переработка информации, проведение ее к другим клеткам. Нейроны выполняют также трофическую функцию, влияя на обмен веществ в аксонах и дендритах. Различают следующие виды нейронов: афферентные, или чувствительные, которые воспринимают раздражение и трансформируют его в нервный импульс; ассоциативные, промежуточные, или интернейроны, которые передают нервный импульс между нейронами; эфферентные, или моторные, которые обеспечивают передачу нервного импульса на рабочую структуру. Эта классификация нейронов основывается на положении нервной клетки в составе рефлекторной дуги. Нервное возбуждение по ней передается лишь в одном направлении. Это правило получило название физиологической, или динамической, поляризации нейронов. Что касается изолированного нейрона, то он способен проводить импульс в любом направлении. Нейроны коры большого мозга по морфологическим признакам делятся на пирамидные и непирамидные.
Нервные клетки контактируют между собой через синапсы специализированные структуры, где нервный импульс переходит из нейрона на нейрон. Большей частью синапсы образуются между аксонами одной клетки и дендритами другой. Различают также другие типы синаптических контактов: аксосоматические, аксоаксональные, дендродентритные. Итак, любая часть нейрона может образовывать синапс с разными частями другого нейрона. Типичный нейрон может иметь от 1000 до 10 000 синапсов и получать информацию от 1000 других нейронов. В составе синапса различают две части -пресинаптическую и постсинаптическую, между которыми находится синаптическая щель. Пресинаптическая часть образована терминальной веточкой аксона той нервной клетки, которая передает импульс. Большей частью она имеет вид небольшой пуговицы и покрыта пресинаптической мембраной. В пресинаптических окончаниях находятся везикулы, или пузырьки, которые содержат так называемые медиаторы. Медиаторами, или нейротрансмит-терами, являются разные биологически активные вещества. В частности, медиатором холинергических синапсов является ацетилхолин, адренергических - норадреналин и адреналин. Постсинаптическая мембрана содержит особый белок рецептор медиатора. На высвобождение нейромедиатора влияют механизмы нейромодуляции. Эту функцию выполняют нейропептиды и нейрогормоны. Синапс обеспечивает односторонность проведения нервного импульса. По функциональным особенностям различают два вида синапсов - возбуждающие, которые способствуют генерации импульсов (деполяризация), и тормозные, которые могут тормозить действие сигналов (гиперполяризация). Нервным клеткам присущ низкий уровень возбуждения.
Испанский нейрогистолог Рамон-и-Кахаль (1852-1934) и итальянский гистолог Камилло Гольджи (1844-1926) за разработку учения о нейроне как о морфологической единице нервной системы были удостоены Нобелевской премии в области медицины и физиологии (1906 г.). Суть разработанной ими нейронной доктрины заключается в следующем.
1. Нейрон является анатомической единицей нервной системы; он состоит из тела нервной клетки (перикарион), ядра нейрона и аксона / дендритов. Тело нейрона и его отростки покрыты цитоплазматической частично проницаемой мембраной, которая выполняет барьерную функцию.
2. Каждый нейрон является генетической единицей, развивается из независимой эмбриональной клетки-нейробласта; генетический код нейрона точно определяет его структуру, метаболизм, связи, которые генетически запрограммированы.
3. Нейрон является функциональной единицей, способной воспринимать стимул, генерировать его и передавать нервный импульс. Нейрон функционирует как единица лишь в коммуникационном звене; в изолированном состоянии нейрон не функционирует. Нервный импульс передается на другую клетку через терминальную структуру - синапс, с помощью нейротранс-миттера, который может тормозить (гиперполяризация) или возбуждать (деполяризация) последующие нейроны на линии. Нейрон генерирует или не генерирует нервный импульс в соответствии с законом «все или ничего».
4. Каждый нейрон проводит нервный импульс лишь в одном направлении: от дендрита к телу нейрона, аксону, синаптическому соединению (динамическая поляризация нейронов).
5. Нейрон является патологической единицей, т. е. реагирует на повреждение как единица; при сильных повреждениях нейрон гибнет как клеточная единица. Процесс дегенерации аксона или миелиновой оболочки дистальнее места повреждения называется валлеровской дегенерацией (перерождением).
6. Каждый нейрон является регенеративной единицей: у человека регенерируют нейроны периферической нервной системы; проводящие пути в пределах центральной нервной системы эффективно не регенерируют.
Таким образом, в соответствии с нейронной доктриной нейрон является анатомической, генетической, функциональной, поляризованной, патологической и регенеративной единицей нервной системы.
Кроме нейронов, которые образовывают паренхиму нервной ткани, важным классом клеток центральной нервной системы являются глиальные клетки (астроциты, олигодендроциты и микроглиоциты), количество которых в 10-15 раз превышает количество нейронов и которые формируют нейроглию. Ее функции: опорная, разграничительная, трофическая, секреторная, защитная. Глиальные клетки принимают участие в высшей нервной (психической) деятельности. При их участии осуществляется синтез медиаторов центральной нервной системы. Нейроглия играет важную роль также в синаптической передаче. Она обеспечивает структурную и метаболическую защиту для сетки нейронов. Итак, между нейронами и глиальными клетками существуют разнообразные морфофункциональные связи.
РАЗВИТИЕ НЕРВНОЙ СИСТЕМЫ ЧЕЛОВЕКА
ФОРМИРОВАНИЕ МОЗГА ОТ МОМЕНТА ОПЛОДОТВОРЕНИЯ ДО РОЖДЕНИЯ
После слияния яйцеклетки со сперматозоидом (оплодотворения) новая клетка начинает делиться. Через некоторое время из этих новых клеток образуется пузырек. Одна стенка пузырька впячивается внутрь, и в результате образуется зародыш, состоящий из трех слоев клеток: самый внешний слой – эктодерма, внутренний – эндодерма и между ними – мезодерма. Нервная система развивается из наружного зародышевого листка – эктодермы. У человека в конце 2-й недели после оплодотворения обособляется участок первичного эпителия и образуется нервная пластинка. Ее клетки начинают делиться и дифференцироваться, вследствие чего они резко отличаются от соседних клеток покровного эпителия (рис. 1.1). В результате деления клеток края нервной пластинки приподнимаются и появляются нервные валики.
В конце 3-й недели беременности края валиков смыкаются, образуя нервную трубку, которая постепенно погружается в мезодерму зародыша. На концах трубки сохраняются два нейропора (отверстия) – передний и задний. К концу 4-й недели нейропоры зарастают. Головной конец нервной трубки расширяется, и из него начинает развиваться головной мозг, а из оставшейся части – спинной мозг. На этой стадии головной мозг представлен тремя пузырями. Уже на 3–4-й неделе выделяются две области нервной трубки: дорсальная (крыловидная пластинка) и вентральная (базальная пластинка). Из крыловидной пластинки развиваются чувствительные и ассоциативные элементы нервной системы, из базальной – моторные. Структуры переднего мозга у человека целиком развиваются из крыловидной пластинки.
В течение первых 2 мес. беременности образуется основной (среднемозговой) изгиб головного мозга: передний мозг и промежуточный мозг загибаются вперед и вниз под прямым углом к продольной оси нервной трубки. Позже формируются еще два изгиба: шейный и мостовой. В этот же период первый и третий мозговые пузыри разделяются дополнительными бороздами на вторичные пузыри, при этом появляется 5 мозговых пузырей. Из первого пузыря образуются большие полушария головного мозга, из второго – промежуточный мозг, который в процессе развития дифференцируется на таламус и гипоталамус. Из оставшихся пузырей формируются мозговой ствол и мозжечок. В течение 5–10-й недели развития начинается рост и дифференцировка конечного мозга: образуются кора и подкорковые структуры. На этой стадии развития появляются мозговые оболочки, формируются ганглии нервной периферической вегетативной системы, вещество коры надпочечников. Спинной мозг приобретает окончательное строение.
В следующие 10–20 нед. беременности завершается формирование всех отделов головного мозга, идет процесс дифференцировки мозговых структур, который заканчивается только с наступлением половозрелости (рис. 1.2). Полушария становятся самой большой частью головного мозга. Выделяются основные доли (лобная, теменная, височная и затылочная), образуются извилины и борозды больших полушарий. В спинном мозге в шейном и поясничном отделах формируются утолщения, связанные с иннервацией соответствующих поясов конечностей. Окончательный вид приобретает мозжечок. В последние месяцы беременности начинается миелинизация (покрытие нервных волокон специальными чехлами) нервных волокон, которая заканчивается уже после рождения.

Головной и спинной мозг покрыты тремя оболочками: твердой, паутинной и мягкой. Головной мозг заключен в черепную коробку, а спинной мозг – в позвоночный канал. Соответствующие нервы (спинномозговые и черепные) покидают ЦНС через специальные отверстия в костях.
В процессе эмбрионального развития головного мозга полости мозговых пузырей видоизменяются и превращаются в систему мозговых желудочков, которые сохраняют связь с полостью спинномозгового канала. Центральные полости больших полушарий головного мозга образуют боковые желудочки довольно сложной формы. Их парные части имеют в своем составе передние рога, которые находятся в лобных долях, задние рога, находящиеся в затылочных долях, и нижние рога, расположенные в височных долях. Боковые желудочки соединяются с полостью промежуточного мозга, которая является III желудочком. Через специальный проток (сильвиев водопровод) III желудочек соединяется с IV желудочком; IV желудочек образует полость заднего мозга и переходит в спинномозговой канал. На боковых стенках IV желудочка находятся отверстия Люшки, а на верхней стенке – отверстие Мажанди. Благодаря этим отверстиям полость желудочков сообщается с подпаутинным пространством. Жидкость, заполняющая желудочки головного мозга, называется эндолимфой и образуется из крови. Процесс образования эндолимфы протекает в специальных сплетениях кровеносных сосудов, (они называются хороидальными сплетениями). Такие сплетения находятся в полостях III и IV мозговых желудочков.
Сосуды головного мозга. Головной мозг человека очень интенсивно снабжается кровью. Это связано, прежде всего, с тем, что нервная ткань одна из наиболее работоспособных в нашем организме. Даже ночью, когда мы отдыхаем от дневной работы, наш мозг продолжает интенсивно работать (подробнее см. раздел «Активирующие системы мозга»). Кровоснабжение головного мозга происходит по следующей схеме. Головной мозг снабжается кровью по двум парам основных кровеносных сосудов: общим сонным артериям, которые проходят в области шеи и их пульсация легко прощупывается, и паре позвоночных артерий, заключенных в латеральных частях позвоночного столба (см. приложение 2). После того как позвоночные артерии покидают шейный последний позвонок, они сливаются в одну базальную артерию, которая проходит в специальной ложбине на основании моста. На основании мозга в результате слияния перечисленных артерий образуется кольцевой кровеносный сосуд. От него кровеносные сосуды (артерии) веерообразно охватывают весь мозг, включая большие полушария.
Венозная кровь собирается в специальные лакуны и покидает пределы головного мозга по яремным венам. Кровеносные сосуды головного мозга вмонтированы в мягкую мозговую оболочку. Сосуды многократно ветвятся и в виде тонких капилляров проникают в мозговую ткань.
Головной мозг человека надежно защищен от проникновения инфекций так называемым гематоэнцефалическим барьером. Этот барьер формируется уже в первую треть срока беременности и включает в себя три мозговые оболочки (самая внешняя – твердая, затем паутинная и мягкая, которая прилежит к поверхности мозга, в ней находятся кровеносные сосуды) и стенки кровеносных капилляров мозга. Другой составляющей частью этого барьера являются глобальные оболочки вокруг кровеносных сосудов, образованные отростками клеток глии. Отдельные мембраны клеток глии тесно прилегают друг к другу, создавая щелевые контакты между собой.
В головном мозге есть участки, где гематоэнцефалический барьер отсутствует. Это район гипоталамуса, полость III желудочка (субфорникальный орган) и полость IV желудочка (area postrema). Здесь стенки кровеносных сосудов имеют специальные места (так называемый фенестрированный, т.е. продырявленный, эпителий сосудов), в которых из нейронов головного мозга в кровеносное русло выбрасываются гормоны и их предшественники. Подробнее эти процессы будут рассмотрены в гл. 5.
Таким образом, с момента зачатия (слияние яйцеклетки со сперматозоидом) начинается развитие ребенка. За это время, которое занимает почти два десятка лет, развитие человека проходит несколько этапов (табл. 1.1).

Вопросы
1. Этапы развития центральной нервной системы человека.
2. Периоды развития нервной системы ребенка.
3. Что составляет гематоэнцефалический барьер?
4. Из какой части нервной трубки развиваются сенсорные и моторные элементы центральной нервной системы?
5. Схема кровоснабжения головного мозга.
Литература
Коновалов А. Н., Блинков С. М., Пуцило М. В. Атлас нейрохирургической анатомии. М., 1990.
Моренков Э. Д. Морфология мозга человека. М.: Изд-во Моск. ун-та, 1978.
Оленев С. Н. Развивающийся мозг. Л., 1979.
Савельев С. Д. Стереоскопический атлас мозга человека. М.: Area XVII, 1996.
Шаде Дж., Форд П. Основы неврологии. М., 1976.
Этапы развития центральной нервной системы
Появление многоклеточных организмов явилось первичным стимулом для дифференциации систем связи, которые обеспечивают целостность реакций организма, взаимодействие между его тканями и органами. Это взаимодействие может осуществляться как гуморальным путем посредством поступления гормонов и продуктов метаболизма в кровь, лимфу и тканевую жидкость, так и за счет функции нервной системы, которая обеспечивает быструю передачу возбуждения, адресованного к вполне определенным мишеням.
Нервная система беспозвоночных животных
Нервная система как специализированная система интеграции на пути структурного и функционального развития проходит через несколько этапов, которые у первично- и вторичноротых животных могут характеризоваться чертами параллелизма и филогенетической пластичностью выбора.
Среди беспозвоночных наиболее примитивный тип нервной системы в виде диффузной нервной сети встречается у типа кишеч-нополостных. Их нервная сеть представляет собой скопление муль-типолярных и биполярных нейронов, отростки которых могут перекрещиваться, прилегать друг к другу и лишены функциональной дифференциации на аксоны и дендриты. Диффузная нервная сеть не разделена на центральный и периферический отделы и может быть локализована в эктодерме и энтодерме.
Эпидермальные нервные сплетения, напоминающие нервные сети кишечнополостных, могут быть обнаружены и у более высоко организованных беспозвоночных (плоские и кольчатые черви), однако здесь они занимают соподчиненное положение по отношению к центральной нервной системе (ЦНС), которая выделяется как самостоятельный отдел.
В качестве примера такой централизации и концентрации нервных элементов можно привести ортогональную нервную систему плоских червей. Ортогон высших турбеллярий представляет собой упорядоченную структуру, которая состоит из ассоциативных и двигательных клеток, формирующих вместе несколько пар продольных тяжей, или стволов, соединенных большим числом поперечных и кольцевых комиссуральных стволов. Концентрация нервных элементов сопровождается их погружением в глубь тела.
Плоские черви являются билатерально симметричными животными с четко выраженной продольной осью тела. Движение у свободноживущих форм осуществляется преимущественно в сторону головного конца, где концентрируются рецепторы, сигнализирующие о приближении источника раздражения. К числу таких рецепторов турбеллярий относятся пигментные глазки, обонятельные ямки, ста-тоцист, чувствительные клетки покровов, наличие которых способствует концентрации нервной ткани на переднем конце тела. Этот процесс приводит к формированию головного ганглия, который, по меткому выражению Ч. Шеррингтона, можно рассматривать как ганглиозную надстройку над системами рецепции на расстоянии.
Ганглионизация нервных элементов получает дальнейшее развитие у высших беспозвоночных, кольчатых червей, моллюсков и членистоногих. У большинства кольчатых червей брюшные стволы ганглионизированы таким образом, что в каждом сегменте тела формируется по одной паре ганглиев, соединенных коннективами с другой парой, расположенной в соседнем сегменте.
Ганглии одного сегмента у примитивных аннелид соединены между собой поперечными комиссурами, и это приводит к образованию лестничной нервной системы,. В более продвинутых отрядах кольчатых червей наблюдается тенденция к сближению брюшных стволов вплоть до полного слияния ганглиев правой и левой сторон и перехода от лестничной к цепочечной нервной системе. Идентичный, цепочечный тип строения нервной системы существует и у членистоногих с различной выраженностью концентрации нервных элементов, которая может осуществляться не только за счет слияния соседних ганглиев одного сегмента, но и при слиянии последовательных ганглиев различных сегментов.
Эволюция нервной системы беспозвоночных идет не только по пути концентрации нервных элементов, но и в направлении усложнения структурных взаимоотношений в пределах ганглиев. Не случайно в современной литературе отмечается тенденция сравнивать брюшную нервную цепочку со спинным мозгом позвоночных животных. Как и в спинном мозгу, в ганглиях обнаруживается поверхностное расположение проводящих путей, дифференциация нейропиля на моторную, чувствительную и ассоциативные области. Это сходство, являющееся примером параллелизма в эволюции тканевых структур, не исключает, однако, своеобразия анатомической организации. Так, например, расположение туловищного мозга кольчатых червей и членистоногих на брюшной стороне тела обусловило локализацию моторного нейропиля на дорсальной стороне ганглия, а не на вентральной, как это имеет место у позвоночных животных.
Процесс ганглионизации у беспозвоночных может привести к формированию нервной системы разбросанно-узлового типа, которая встречается у моллюсков. В пределах этого многочисленного типа имеются филогенетически примитивные формы с нервной системой, сопоставимой с ортогоном плоских червей (боконервные моллюски), и продвинутые классы (головоногие моллюски), у которых слившиеся ганглии формируют дифференцированный на отделы мозг.
Прогрессивное развитие мозга у головоногих моллюсков и насекомых создает предпосылку для возникновения своеобразной иерархии командных систем управления поведением. Низший уровень интеграции в сегментарных ганглиях насекомых и в подглоточной массе мозга моллюсков служит основой для автономной деятельности и координации элементарных двигательных актов. В то же время мозг представляет собой следующий, более высокий уровень интеграции, где могут осуществляться межанализаторный синтез и оценка биологической значимости информации. На основе этих процессов формируются нисходящие команды, обеспечивающие вариантность запуска нейронов сегментарных центров. Очевидно, взаимодействие двух уровней интеграции лежит в основе пластичности поведения высших беспозвоночных, включающего врожденные и приобретенные реакции.
В целом, говоря об эволюции нервной системы беспозвоночных, было бы упрощением представлять ее как линейный процесс. Факты, полученные в нейроонтогенетических исследованиях беспозвоночных, позволяют допустить множественное (полигенетическое) происхождение нервной ткани беспозвоночных. Следовательно, эволюция нервной системы беспозвоночных могла идти широким фронтом от нескольких источников с изначальным многообразием.
На ранних этапах филогенетического развития сформировался второй ствол эволюционного древа, который дал начало иглокожим и хордовым. Основным критерием для выделения типа хордовых является наличие хорды, глоточных жаберных щелей и дорсального нервного тяжа - нервной трубки, представляющей собой производное наружного зародышевого листка - эктодермы. Трубчатый тип нервной системы позвоночных по основным принципам организации отличен от ганглионарного или узлового типа нервной системы высших беспозвоночных.
Нервная система позвоночных животных
Нервная система позвоночных закладывается в виде сплошной нервной трубки, которая в процессе онто- и филогенеза дифференцируется на различные отделы и является также источником периферических симпатических и парасимпатических нервных узлов. У наиболее древних хордовых (бесчерепных) головной мозг отсутствует и нервная трубка представлена в малодифференцированном состоянии.
Согласно представлениям Л. А. Орбели, С. Херрика, А. И. Ка-рамяна, этот критический этап развития центральной нервной си-
стемы обозначается как спинальный. Нервная трубка современного бесчерепного (ланцетника), как и спинной мозг более высоко организованных позвоночных, имеет метамерное строение и состоит из 62-64 сегментов, в центре которых проходит спинно-мозговой канал. От каждого сегмента отходят брюшные (двигательные) и спинные (чувствительные) корешки, которые не образуют смешанных нервов, а идут в виде отдельных стволов. В головных и хвостовых отделах нервной трубки локализованы гигантские клетки Родэ, толстые аксоны которых образуют проводниковый аппарат. С клетками Родэ связаны светочувствительные глазки Гесса, возбуждение которых вызывает отрицательный фототаксис.
В головной части нервной трубки ланцетника находятся крупные ганглиозные клетки Овсянникова, имеющие синаптические контакты с биполярными чувствительными клетками обонятельной ямки. В последнее время в головной части нервной трубки идентифицированы нейросекреторные клетки, напоминающие гипофи-зарную систему высших позвоночных. Однако анализ восприятия и простых форм обучения ланцетника показывает, что на данном этапе развития ЦНС функционирует по принципу эквипотенци-альности, и утверждение о специфике головного отдела нервной трубки не имеет достаточных оснований.
В ходе дальнейшей эволюции наблюдается перемещение некоторых функций и систем интеграции из спинного мозга в головной - процесс энцефализации, который был рассмотрен на примере беспозвоночных животных. В период филогенетического развития от уровня бесчерепных до уровня круглоротых формируется головной мозг как надстройка над системами дистантной рецепции.
Исследование ЦНС современных круглоротых показывает, что их головной мозг в зачаточном состоянии содержит все основные структурные элементы. Развитие вестибулолатеральной системы, связанной с полукружными каналами и рецепторами боковой линии, возникновение ядер блуждающего нерва и дыхательного центра создают основу для формирования заднего мозга. Задний мозг миноги включает продолговатый мозг и мозжечок в виде небольших выпячиваний нервной трубки.
Развитие дистантной зрительной рецепции дает толчок к закладке среднего мозга. На дорсальной поверхности нервной трубки развивается зрительный рефлекторный центр - крыша среднего мозга, куда приходят волокна зрительного нерва. И наконец, развитие обонятельных рецепторов способствует формированию переднего или конечного мозга, к которому примыкает слаборазвитый промежуточный мозг.
Указанная выше направленность процесса энцефализации согласуется с ходом онтогенетического развития мозга у круглоротых. В процессе эмбриогенеза головные отделы нервной трубки дают начало трем мозговым пузырям. Из переднего пузыря формируется конечный и промежуточный мозг, средний пузырь дифференцируется в средний мозг, а из заднего пузыря образуются продолговатый
мозг и мозжечок. Сходный план онтогенетического развития мозга сохраняется и у других классов позвоночных.
Нейрофизиологические исследования мозга круглоротых показывают, что его главный интегративный уровень сосредоточен в среднем и продолговатом мозгу, т. е. на данном этапе развития ЦНС доминирует бульбомезенцефальная система интеграции, пришедшая на смену спинальной.
Передний мозг круглоротых длительное время считали чисто обонятельным. Однако исследования недавнего времени показали, что обонятельные входы в передний мозг не являются единственными, а дополняются сенсорными входами других модальностей. Очевидно, уже на ранних этапах филогенеза позвоночных передний мозг начинает участвовать в переработке информации и управлении поведением.
Вместе с тем энцефализация как магистральное направление развития мозга не исключает эволюционных преобразований в спинном мозгу круглоротых. В отличие от бесчерепных нейроны кожной чувствительности выделяются из спинного мозга и концентрируются в спинно-мозговой ганглий. Наблюдается совершенствование проводниковой части спинного мозга. Проводящие волокна боковых столбов имеют контакты с мощной дендритной сетью мотонейронов. Формируются нисходящие связи головного мозга со спинным через мюллеровские волокна - гигантские аксоны клеток, лежащих в среднем и продолговатом мозгу.
Появление более сложных форм двигательного поведения у позвоночных сопряжено с совершенствованием организации спинного мозга. Так, например, переход от стереотипных ундулирующих движений круглоротых к локомоции с помощью плавников у хрящевых рыб (акулы, скаты) связан с разделением кожной и мышеч-но-суставной (проприоцептивной) чувствительности. В спинальных ганглиях появляются специализированные нейроны для выполнения этих функций.
В эфферентной части спинного мозга хрящевых рыб также наблюдаются прогрессивные преобразования. Укорачивается путь моторных аксонов внутри спинного мозга, происходит дальнейшая дифференциация его проводящих путей. Восходящие пути боковых столбов у хрящевых рыб доходят до продолговатого мозга и мозжечка. Вместе с тем восходящие пути задних столбов спинного мозга еще не дифференцированы и состоят из коротких звеньев.
Нисходящие пути спинного мозга у хрящевых рыб представлены развитым ретикулоспинальным трактом и путями, соединяющими вестибулолатеральную систему и мозжечок со спинным мозгом (вестибулоспинальный и церебеллоспинальный тракты).
Одновременно в продолговатом мозгу наблюдается усложнение системыядер вестибулолатеральной зоны. Этот процесс сопряжен с дальнейшей дифференциацией органов боковой линии и с появлением в лабиринте третьего (наружного) полукружного канала в Дополнение к переднему и заднему.
Развитие общей двигательной координации у хрящевых рыб связано с интенсивным развитием мозжечка. Массивный мозжечок акулы имеет двусторонние связи со спинным, продолговатым мозгом и покрышкой среднего мозга. Функционально он разделяется на две части: старый мозжечок (архицеребеллум), связанный с вестибуло-латеральной системой, и древний мозжечок (палецоцеребеллум), включенный в систему анализа проприоцептивной чувствительности. Существенным моментом структурной организации мозжечка хрящевых рыб является его многослойность. В сером веществе мозжечка акулы идентифицированы молекулярный слой, слой клеток Пуркинье и зернистый слой.
Другой многослойной структурой стволовой части мозга хрящевых рыб является крыша среднего мозга, куда подходят афференты различных модальностей (зрительные, соматические). Сама морфологическая организация среднего мозга свидетельствует о его важной роли в интегративных процессах на данном уровне филогенетического развития.
В промежуточном мозгу хрящевых рыб происходит дифференциация гипоталамуса, который является наиболее древним образованием этой части мозга. Гипоталамус имеет связи с конечным мозгом. Сам конечный мозг разрастается и состоит из обонятельных луковиц и парных полушарий. В полушариях у акул находятся зачатки старой коры (архикортекса) и древней коры (палеокортекса).
Палеокортекс, тесно связанный с обонятельными луковицами, служит главным образом для восприятия обонятельных стимулов. Архикортекс, или гиппокампальная кора, предназначен для более сложной обработки обонятельной информации. Вместе с тем электрофизиологические исследования показали, что обонятельные проекции занимают только часть полушарий переднего мозга акул. Кроме обонятельной здесь обнаружено представительство зрительной и соматической сенсорных систем. Очевидно, старая и древняя кора может участвовать в регуляции поисковых, пищевых, половых и оборонительных рефлексов у хрящевых рыб, многие из которых являются активными хищниками.
Таким образом, у хрящевых рыб складываются основные черты ихтиопсидного типа организации мозга. Его отличительной чертой является присутствие надсегментарного аппарата интеграции, координирующего работу моторных центров и организующего поведение. Эти интегративные функции осуществляют средний мозг и мозжечок, что позволяет говорить о мезэнцвфалоцеребеллярной системе интеграции на данном этапе филогенетического развития нервной системы. Конечный мозг остется преимущественно обонятельным, хотя и участвует в регуляции функций нижележащих отделов.
Переход позвоночных от водного к наземному образу жизни связан с целым рядом перестроек в ЦНС. Так, например, у амфибий в спинном мозгу появляется два утолщения, соответствующие верхнему и нижнему поясам конечностей. В спинальных ганглиях вместо биполярных чувствительных нейронов сосредоточиваются униполярные с Т-образно ветвящимся отростком, обеспечивающим более высокую скорость проведения возбуждения без участия клеточного тела. На периферии в коже земноводных формируются специализированные рецепторы и рецепторные поля, обеспечивающие дискриминационную чувствительность.
В мозговом стволе также происходят структурные изменения в связи с перераспределением функциональной значимости различных отделов. В продолговатом мозгу наблюдаются редукция ядер боковой линии и формирование кохлеарного, слухового ядра, осуществляющего анализ информации от примитивного органа слуха.
По сравнению с рыбами у амфибий, имеющих довольно стереотипную локомоцию, наблюдается значительная редукция мозжечка. Средний мозг, так же как и у рыб, представляет собой многослойную структуру, в которой наряду с передним двухолмием - ведущим отделом интеграции зрительного анализатора - появляются дополнительные бугорки - предшественники задних холмов четверохолмия.
Наиболее существенные в эволюционном плане изменения происходят в промежуточном мозгу амфибий. Здесь обособляется зрительный бугор - таламус, появляются структурированные ядра (наружное коленчатое тело) и восходящие пути, связывающие зрительный бугор с корой (таламокортикальный тракт).
В полушариях переднего мозга происходит дальнейшая дифференциация старой и древней коры. В старой коре (архикортексе) обнаруживаются звездчатые и пирамидные клетки. В промежутке между старой и древней корой появляется полоска плаща, которая является предтечей новой коры (неокортекса).
В целом развитие переднего мозга создает предпосылки для перехода от свойственной рыбам мозжечково-мезэнцефальной системы интеграции к диэнцефало-телэнцефальной, где ведущим отделом становится передний мозг, а зрительный бугор промежуточного мозга превращается в коллектор всех афферентных сигналов. В полной мере эта система интеграции представлена в зауропсидном типе мозга у рептилий и знаменует собой следующий этап мор-фофункциональной эволюции мозга .
Развитие таламокортикальной системы связей у рептилий приводит к формированию новых проводящих путей, как бы подтягивающихся к филогенетически молодым формациям мозга.
В боковых столбах спинного мозга рептилий появляется восходящий спиноталамический тракт, который проводит к головному мозгу информацию о температурной и болевой чувствительности. Здесь же в боковых столбах формируется новый нисходящий тракт - руброспинальный (Монакова). Он связывает мотонейроны спинного мозга с красным ядром среднего мозга, которое включено в древнюю экстрапирамидную систему двигательной регуляции. Эта многозвенная система объединяет влияние переднего мозга, мозжечка, ретикулярной формации ствола, ядер вестибулярного комплекса и координирует двигательную активность.
У рептилий, как истинно наземных животных, возрастает роль зрительной и акустической информации, возникает необходимость
сопоставления этой информации с обонятельной и вкусовой, В соответствии с этими биологическими изменениями в стволовой части мозга рептилий происходит целый ряд структурных изменений. В продолговатом мозгу дифференцируются слуховые ядра, помимо кохлеарного ядра появляется угловое, связанное со средним мозгом. В среднем мозгу двухолмие преобразуется в четверохолмие, в задних холмах которого локализованы акустические центры.
Наблюдается дальнейшая дифференциация связей крыши среднего мозга со зрительным бугром - таламусом, который является как бы преддверием перед входом в кору всех восходящих сенсорных путей. В самом таламусе происходит дальнейшее обособление ядерных структур и установление между ними специализированных связей.
Конечный мозг рептилий может иметь два типа организации:
кортикальный и стриатальный. Кортикальный тип организации, свойственный современным черепахам, характеризуется преимущественным развитием полушарий переднего мозга и параллельным ‘развитием новых отделов мозжечка. В дальнейшем это направление в эволюции мозга сохраняется у млекопитающих.
Стриатальный тип организации, характерный для современных ящериц, отличается доминирующим развитием находящихся в глубине полушарий базальных ганглиев, в частности полосатого тела. По этому пути далее идет развитие головного мозга у птиц. Представляет интерес, что в полосатом теле у птиц имеются клеточные объединения или ассоциации нейронов (от трех до десяти), разделенные олигодендроглией. Нейроны таких ассоциаций получают одинаковую афферентацию, и это делает их сходными с нейронами, объединенными в вертикальные колонки в новой коре млекопитающих. В то же время в полосатом теле млекопитающих идентичные клеточные ассоциации не описаны. Очевидно, это является примером конвергентной эволюции, когда сходные образования развились независимо у различных животных.
У млекопитающих развитие переднего мозга сопровождалось бурным ростом новой коры, находящейся в тесной функциональной связи со зрительным бугром промежуточного мозга. В коре закладываются эфферентные пирамидные клетки, посылающие свои длинные аксоны к мотонейронам спинного мозга.
Таким образом, наряду с многозвенной экстрапирамидной системой появляются прямые пирамидные пути, которые обеспечивают непосредственный контроль над двигательными актами. Корковая регуляция моторики у млекопитающих приводит к развитию филогенетически наиболее молодой части мозжечка - передней части задних долей полушарий, или неоцеребеллума. Неоцеребеллум приобретает двусторонние связи с новой корой.
Рост новой коры у млекопитающих происходит настолько интенсивно, что старая и древняя кора оттесняется в медиальном направлении к мозговой перегородке. Бурный рост коры компенсируется формированием складчатости. У наиболее низко организованных однопроходных (утконос) на поверхности полушария закладываются первые две постоянные борозды, остальная же поверхность остается гладкой (лиссэнцефальный тип коры).
Как показали нейрофизиологические исследования, мозг однопроходных и сумчатых млекопитающих лишен еще соединяющего полушария мозолистого тела и характеризуется перекрытием сенсорных проекций в новой коре. Четкая локализация моторных, зрительных и слуховых проекций здесь отсутствует.
У плацентарных млекопитающих (насекомоядных и грызунов) * отмечается развитие более четкой локализации проекционных зон в коре. Наряду с проекционными зонами в новой коре формируются ассоциативные зоны, однако границы первых и вторых могут перекрываться. Мозг насекомоядных и грызунов характеризуется наличием мозолистого тела и дальнейшим увеличением общей площади новой коры.
В процессе параллельно-адаптивной эволюции у хищных млекопитающих появляются теменные и лобные ассоциативные поля, ответственные за оценку биологически значимой информации, мотивацию поведения и программирование сложных поведенческих актов. Наблюдается дальнейшее развитие складчатости новой коры.
И наконец, приматы демонстрируют наиболее высокий уровень организации коры головного мозга. Кора приматов характеризуется шестислойностью, отсутствием перекрытия ассоциативных и проекционных зон. У приматов формируются связи между фронтальными и теменными ассоциативными полями и, таким образом, возникает целостная интегративная система больших полушарий.
В целом, прослеживая основные этапы эволюции мозга позвоночных, следует отметить, что его развитие не сводилось просто к линейному увеличению размеров. В различных эволюционных линиях позвоночных могли иметь место независимые процессы увеличения размеров и усложнения цитоархитектоники различных отделов мозга. Примером тому может служить сравнение стриа-тального и кортикального типов организации переднего мозга позвоночных.
В процессе развития наблюдается тенденция к перемещению ведущих интегративных центров мозга в ростральном направлении от среднего мозга и мозжечка к переднему мозгу. Однако эту тенденцию нельзя абсолютизировать, так как мозг представляет собой целостную систему, в которой стволовые части играют важную функциональную роль на всех этапах филогенетического развития позвоночных. Кроме того, начиная с круглоротых в переднем мозгу обнаруживаются проекции различных сенсорных модальностей, свидетельствующие об участии этого отдела мозга в управлении поведением уже на ранних стадиях эволюции позвоночных.
Список литературы
1. Самусев Р.П. Анатомия человека.М., 1995.
2. Анатомия человека/ Под ред. М.Р. Сапина. М.,1986.
3. Общий курс физиологии человека и животных в 2-х кн. Под ред. А.Д.Ноздрачёва. М., “Высшая школа”,1991.
В эволюции нервная система претерпела несколько этапов развития, которые стали поворотными пунктами в качественной организации её деятельности. Эти этапы отличаются по количеству и видам нейрональных образований, синапсов, признакам их функциональной специализации, по образованию группировок нейронов, связанных между собой общностью функций. Выделяют три основных этапа структурной организации нервной системы: диффузный, узловой, трубчатый.
Диффузная нервная система наиболее древняя, имеется у кишечнополостных (гидра) животных. Такая нервная система характеризуется множественностью связей соседних элементов, что позволяет возбуждению свободно распространяться по нервной сети во все стороны.
Этот тип нервной системы обеспечивает широкую взаимозаменяемость и тем самым большую надёжность функционирования, однако эти реакции имеют неточный, расплывчатый характер.
Узловой тип нервной системы типичен для червей, моллюсков, ракообразных.
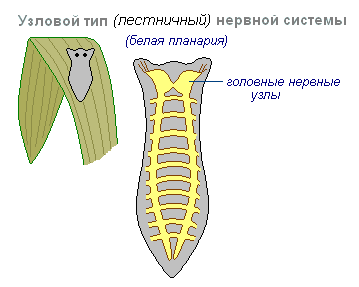
Он характерен тем, что связи нервных клеток организованы определённым образом, возбуждение проходит по жёстко определённым путям. Такая организация нервной системы оказывается более ранимой. Повреждение одного узла вызывает нарушение функций всего организма в целом, но она по своим качествам быстрее и точнее.
Трубчатая нервная система характерна для хордовых, она включает в себя черты диффузного и узлового типов. Нервная система высших животных взяла всё лучшее: высокую надёжность диффузного типа, точность, локальность быстроту организации реакций узлового типа.
Ведущая роль нервной системы
На первом этапе развития мира живых существ взаимодействие между простейшими организмами осуществлялось через водную среду первобытного океана, в которую поступали химические вещества, выделяемые ими. Первой древнейшей формой взаимодействия между клетками многоклеточных организм является химическое взаимодействие посредством продуктов обмена веществ, поступающих в жидкости организма. Такими продуктами обмена веществ, или метаболитами, являются продукты распада белков, углекислота и др. это — гуморальная передача влияний, гуморальный механизм корреляции, или связи между органами.
Гуморальная связь характеризуется следующими особенностями:
- отсутствием точного адреса, по которому направляется химическое вещество, поступающее в кровь или другие жидкости тела;
- химическое вещество распространяется медленно;
- химическое вещество действует в ничтожных количествах и обычно быстро разрушается или выводится из организма.
Гуморальные связи являются общими и для мира животных, и для мира растений. На определённой ступени развития мира животных в связи с появлением нервной системы образуется новая, нервная форма связей и регуляций, которая качественно отличает мир животных от мира растений. Чем выше по своему развитию организм животного, тем большую роль играет взаимодействие органов через нервную систему, которое обозначается как рефлекторное. У высших живых организмов нервная система регулирует гуморальные связи. В отличие от гуморальной связи нервная связь имеет точную направленность к определённому органу и даже группе клеток; связь осуществляется в сотни раз с большей скоростью, чем скорость распространения химических веществ. Переход от гуморальной связи к нервной сопровождался не уничтожением гуморальной связи между клетками тела, а подчинением нервным связям и возникновению нервно-гуморальным связям.
На следующем этапе развития живых существ появляются специальные органы — железы, в которых вырабатываются гормоны, образующиеся из поступающих в организм пищевых веществ. Основная функция нервной системы заключается как в регуляции деятельности отдельных органов между собой, так и во взаимодействии организма как единого целого с окружающей его внешней средой. Любое воздействие внешней среды на организм оказывается, прежде всего, на рецепторы (органы чувств) и осуществляется через посредство изменений, вызываемых внешней средой и нервной системой. По мере развития нервной системы высший её отдел — большие полушария головного мозга — становится «распорядителем и распределителем всей деятельности организма».
Строение нервной системы
Нервная система образована нервной тканью, которая состоит из огромного количества нейронов — нервная клетка с отростками.

Нервная система условно подразделяется на центральную и периферическую.

Центральная нервная система включает головной и спинной мозг, а периферическая нервная система - нервы, отходящие от них.

Головной и спинной мозг представляют собой совокупность нейронов. На поперечном разрезе мозга различают белое и серое вещество. Серое вещество состоит из нервных клеток, а белое - из нервных волокон, являющихся отростками нервных клеток. В различных отделах центральной нервной системы расположение белого и серого вещества неодинаково. В спинном мозге серое вещество находится внутри, а белое — снаружи, в головном же (большие полушария, мозжечок), наоборот — серое вещество — снаружи, белое — внутри. В различных отделах головного мозга имеются отдельные скопления нервных клеток (серого вещества), расположенные внутри белого вещества, - ядра . Скопления нервных клеток находятся и за пределами центральной нервной системы. Они называются узлами и относятся к периферической нервной системе.
Рефлекторная деятельность нервной системы
Основной формой деятельности нервной системы является рефлекс. Рефлекс - реакция организма на изменение внутренней или внешней среды, осуществляемая при участии центральной нервной системы в ответ на раздражение рецепторов.

При всяком раздражении возбуждение с рецепторов передаётся по центростремительным нервным волокнам в центральную нервную систему, откуда через вставочный нейрон по центробежным волокнам оно идёт на периферию к тому или иному органу, деятельность которого изменяется. Весь этот путь через центральную нервную систему к рабочему органу, называется рефлекторной дугой образован обычно тремя нейронами: чувствительным, вставочным и двигательным. Рефлекс — сложный акт, в осуществлении которого принимает участие значительно большее количество нейронов. Возбуждение, попадая в центральную нервную систему, распространяется на многие отделы спинного мозга и доходит до головного. В результате взаимодействия многих нейронов осуществляется ответная реакция организма на раздражение.
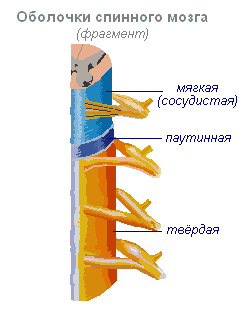
Спинной мозг
Спинной мозг - тяж длиной около 45 см, диаметром 1 см, находится в канале позвоночника, покрыт тремя мозговыми оболочками: твёрдой, паутинной и мягкой (сосудистой).
Спинной мозг находится в позвоночном канале и представляет собой тяж, который вверху переходит в продолговатый мозг, а внизу заканчивается на уровне второго поясничного позвонка. Спинной мозг состоит из серого вещества, содержащего нервные клетки, и белого, состоящего из нервных волокон. Серое вещество расположено внутри спинного мозга и окружено со всех сторон белым веществом.
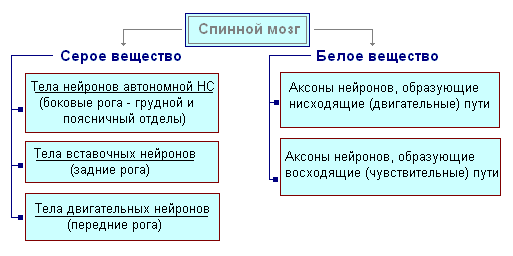
На поперечном разрезе серое вещество напоминает букву Н. В нём различают передние и задние рога, а также соединяющую перекладину, в центре которой находится узкий канал спинного мозга, содержащий спинномозговую жидкость. В грудном отделе выделяют боковые рога. В них заложены тела нейронов, иннервирующих внутренние органы. Белое вещество спинного мозга образовано нервными отростками. Короткие отростки соединяют участки спинного мозга, а длинные составляют проводниковый аппарат двусторонних связей с головным мозгом.
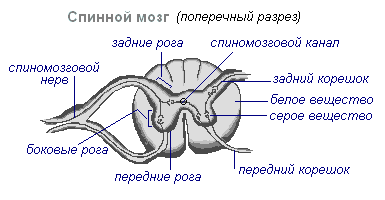
Спинной мозг имеет два утолщения - шейное и поясничное, от которых отходят нервы к верхним и нижним конечностям. От спинного мозга отходит 31 пара спинномозговых нервов. Каждый нерв начинается от спинного мозга двумя корешками — передним и задним. Задние корешки — чувствительные состоят из отростков центростремительных нейронов. Их тела расположены в спинномозговых узлах. Передние корешки — двигательные — являются отростками центробежных нейронов расположенных в сером веществе спинного мозга. В результате слияния переднего и заднего корешка образуется смешанный спинномозговой нерв. В спинном мозге сосредоточены центры, регулирующие наиболее простые рефлекторные акты. Основные функции спинного мозга - рефлекторная деятельность и проведение возбуждения.

В спинном мозге человека заложены рефлекторные центры мышц верхних и нижних конечностей, потоотделения и мочеиспускания. Функции проведения возбуждения заключается в том, что через спинной мозг проходят импульсы от головного мозга ко всем областям тела и обратно. По восходящим проводящим путям в головной мозг передаются центростемительные импульсы от органов (кожа, мышцы). По нисходящим путям центробежные импульсы передаются от головного мозга в спинной, затем на периферию, к органам. При повреждении проводящих путей наблюдается потеря чувствительности в различных участках тела, нарушение произвольных сокращений мышц и способности к движению.
Эволюция головного мозга позвоночных
Образование центральной нервной системы в виде нервной трубки впервые появляется у хордовых. У низших хордовых нервная трубка сохраняется в течение всей жизни, у высших - позвоночных - в стадии эмбриона на спинной стороне закладывается нервная пластинка, которая погружается под кожу и сворачивается в трубку. В эмбриональной стадии развития нервная трубка образует в передней части три вздутия — три мозговых пузыря, из которых развиваются отделы мозга: передний пузырь дает передний и промежуточный мозг, средний пузырь превращается в средний мозг, задний пузырь образует мозжечок и продолговатый мозг . Эти пять отделов мозга характерны для всех позвоночных животных.
Для низших позвоночных - рыб и земноводных - характерно преобладание среднего мозга над остальными отделами. У земноводных несколько увеличивается передний мозг и в крыше полушарий образуется тонкий слой нервных клеток - первичный мозговой свод, древняя кора. У рептилий значительно увеличивается передний мозг за счет скоплений нервных клеток. Большую часть крыши полушарий занимает древняя кора. Впервые у рептилий появляется зачаток новой коры. Полушария переднего мозга наползают на другие отделы, вследствие чего образуется изгиб в области промежуточного мозга. Начиная с древних рептилий, полушария головного мозга становятся самым большим отделом головного мозга.

В строении головного мозгаптиц и пресмыкающихся много общего. На крыше головного мозга - первичная кора, хорошо развит средний мозг. Однако у птиц по сравнению с рептилиями возрастают общая масса мозга и относительные размеры переднего мозга. Мозжечок крупный и имеет складчатое строение. У млекопитающих передний мозг достигает наибольшей величины и сложности. Большую часть мозгового вещества составляет новая кора, которая служит центром высшей нервной деятельности. Промежуточный и средний отделы мозга у млекопитающих невелики. Разрастающиеся полушария переднего мозга накрывают их и подминают под себя. У некоторых млекопитающих мозг гладкий, без борозд и извилин, но у большинства млекопитающих в коре мозга имеются борозды и извилины. Появление борозд и извилин происходит вследствие роста мозга при ограниченных размерах черепа. Дальнейший рост коры приводит к появлению складчатости в виде борозд и извилин.
Головной мозг
Если спинной мозг у всех позвоночных животных развит более или менее одинаково, то головной мозг существенно отличатся размерами и сложностью строения у разных животных. Особенно резкие изменения в ходе эволюции претерпевает передний мозг. У низших позвоночных передний мозг развит слабо. У рыб он представлен обонятельными долями и ядрами серого вещества в толще мозга. Интенсивное развитие переднего мозга связано с выходом животных на сушу. Он дифференцируется на промежуточный мозг и на два симметричных полушария, которые называются конечным мозгом . Серое вещество на поверхности переднего мозга (кора) впервые появляется у пресмыкающихся, развиваясь далее у птиц и особенно у млекопитающих. Действительно большими полушариями переднего мозга становятся только у птиц и млекопитающих. У последних они покрывают почти все другие отделы головного мозга.
Головной мозг расположен в полости черепа. В него входят ствол и конечный мозг (кора больших полушарий).

Ствол мозга состоит из продолговатого мозга, варолиева моста, среднего и промежуточного мозга.
Продолговатый мозг является непосредственным продолжением спинного мозга и расширяясь, переходит в задний мозг. Он в основном сохраняет форму и строение спинного мозга. В толще продолговатого мозга расположены скопления серого вещества — ядра черепно-мозговых нервов. В состав заднего моста входят мозжечок и варолиев мост . Мозжечок расположен над продолговатым мозгом и имеет сложное строение. На поверхности полушарий мозжечка серое вещество образует кору, а внутри мозжечка - его ядра. Как и спинной продолговатый мозг выполняет две функции: рефлекторную и проводниковую. Однако рефлексы продолговатого мозга более сложные. Это выражается в важном значении в регуляции сердечной деятельности, состоянии сосудов, дыхания, потоотделения. В продолговатом мозге расположены центры всех этих функций. Здесь же находятся центры жевания, сосания, глотания, отделения слюны и желудочного сока. Несмотря на малую величину (2,5–3 см), продолговатый мозг представляет собой жизненно важный отдел ЦНС. Повреждение его может стать причиной смерти вследствие прекращения дыхания и деятельности сердца. Проводниковая функция продолговатого мозга и варолиева моста заключается в передаче импульсов из спинного мозга в головной и обратно.
В среднем мозге расположены первичные (подкорковые) центры зрения и слуха, которые осуществляют рефлекторные ориентировочные реакции на световые и звуковые раздражения. Эти реакции выражаются в различных движениях туловища, головы и глаз в сторону раздражителей. Средний мозг состоит из ножек мозга и четверохолмия. Средний мозг регулирует и распределяет тонус (напряжение) скелетных мышц.
Промежуточный мозг состоит из двух отделов - таламус и гипоталамус , каждый из которых состоит из большого числа ядер зрительных бугров и подбугровой области. Через зрительные бугры центростремительные импульсы передаются к коре больших полушарий от всех рецепторов тела. Ни один центростремительный импульс, откуда бы он ни шёл, не может пройти к коре, минуя зрительные бугры. Таким образом, через промежуточный мозг осуществляется связь всех рецепторов с корой больших полушарий. В подбугровой области расположены центры, оказывающие влияние на обмен веществ, терморегуляцию и железы внутренней секреции.
Мозжечок находится позади продолговатого мозга. Он состоит из серого и белого вещества. Однако в отличие от спинного мозга и ствола серое вещество - кора - находится на поверхности мозжечка, а белое вещество расположено внутри, под корой. Мозжечок координирует движения, делает их чёткими и плавными, играет важную роль в сохранении равновесия тела в пространстве, а также оказывает влияние на тонус мышц. При поражении мозжечка у человека наблюдается падение тонуса мышц, расстройство движений и изменение походки, замедляется речь и т.д. Однако через некоторое время движения и мышечный тонус восстанавливаются благодаря тому, что неповреждённые участки центральной нервной системы берут на себя функции мозжечка.
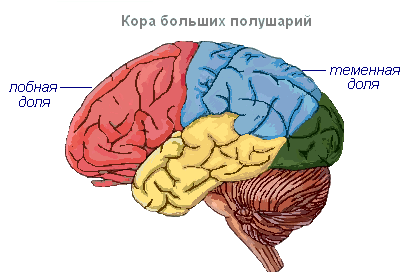
Большие полушария - наиболее крупный и развитый отдел головного мозга. У человека они образуют основную массу головного мозга и по всей своей поверхности покрыты корой. Серое вещество покрывает полушария снаружи и образует кору головного мозга. Кора полушарий человека имеет толщину от 2 до 4 мм и слагается из 6–8 слоёв, образованных 14–16 млрд. клеток, различных по форме, величине и выполняемым функциям. Под корой находится белое вещество. Оно состоит из нервных волокон, связывающих кору с расположенными ниже отделами центральной нервной системы и отдельные доли полушарий между собой.
Кора головного мозга имеет извилины, разделённые бороздами, которые значительно увеличивают её поверхность. Три самые глубокие борозды делят полушария на доли. В каждом полушарии различают четыре доли: лобную, теменную, височную, затылочную . Возбуждение разных рецепторов поступают в соответствующие воспринимающие участки коры, называемые зонами , и отсюда передаются к определённому органу, побуждая его к действию. В коре выделяют следующие зоны. Слуховая зона расположена в височной доле, воспринимает импульсы от слуховых рецепторов.

Зрительная зона лежит в затылочной области. Сюда поступают импульсы от рецепторов глаза.
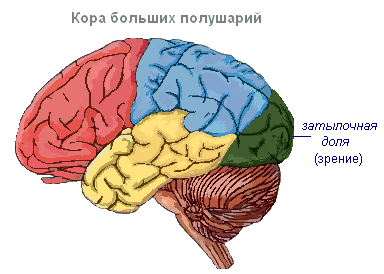
Обонятельная зона находится на внутренней поверхности височной доли и связана с рецепторами носовой полости.

Чувствительно-двигательная зона расположена в лобной и теменной долях. В этой зоне находятся главные центры движения ног, туловища, рук, шеи, языка и губ. Здесь же лежит и центр речи.
Полушария головного мозга - это высший отдел центральной нервной системы, контролирующий работу всех органов у млекопитающих. Значение больших полушарий у человека заключается ещё и в том, что они представляют собой материальную основу психической деятельности. И.П.Павлов показал, что в основе психической деятельности лежат физиологические процессы, происходящие в коре головного мозга. Мышление связано с деятельностью всей коры головного мозга, а не только с функцией отдельных её областей.
| Отдел головного мозга | Функции | |
| Продолговатый мозг | Проводниковая | Связь спинного и вышележащих отделов головного мозга. |
| Рефлекторная | Регуляция деятельности дыхательной, сердечно-сосудистой, пищеварительной систем:
|
|
| Варолиев мост | Проводниковая | Соединяет полушария мозжечка между собой и с корой больших полушарий головного мозга. |
| Мозжечок | Координационная | Координация произвольных движений и сохранение положения тела в пространстве. Регуляция мышечного тонуса и равновесия |
| Средний мозг | Проводниковая | Ориентировочные рефлексы на зрительные, звуковые раздражители (повороты головы и туловища ). |
| Рефлекторная |
|
|
| Промежуточный мозг | таламус
гипоталамус
|
|
Кора больших полушарий
Поверхность коры больших полушарий у человека составляет около 1500 см 2 , что во много раз превышает внутреннюю поверхность черепа. Такая большая поверхность коры образовалась благодаря развитию большого количества борозд и извилин, в результате чего большая часть коры (около 70%) сосредоточена в бороздах. Самые большие борозды больших полушарий - центральная , которая проходит поперёк обоих полушарий, и височная , отделяющая височную долю от остальных. Кора больших полушарий, несмотря на малую толщину (1,5–3 мм) имеет очень сложное строение. В ней насчитывают шесть основных слоёв, которые отличаются строением, формой и размерами нейронов и связями. В коре находятся центры всех чувствительных (рецепторных) систем, представительства всех органов и частей тела. В связи с этим к коре подходят центростремительные нервные импульсы от всех внутренних органов или частей тела, и она может управлять их работой. Через кору больших полушарий происходит замыкание условных рефлексов, посредством которых организм постоянно, в течение всей жизни очень точно приспосабливается к изменчивым условиям существования, к окружающей среде.
- 1)Дорзальная индукция или Первичная нейруляция - период 3-4 недели гестации;
- 2)Вентральная индукция - период 5-6 недели гестации;
- 3)Нейрональная пролиферация - период 2-4 месяца гестации;
- 4)Миграция - период 3-5 месяца гестации;
- 5)Организация - период 6-9 месяца развития плода;
- 6)Миелинизация - занимает период от момента рождения и в последующем периоде постнатальной адаптации.
В первом триместре беременности протекают такие этапы развития нервной системы плода:
Дорзальная индукция или Первичная нейруляция - в связи с индивидуальными особенностями развития может варьировать по времени, но всегда придерживается 3-4 неделе (18-27 день после зачатия) гестации. В этот период происходит образование нервной пластинки, которая после смыкания ее краев превращается в нервную трубку (4-7 неделя гестации).
Вентральная индукция - этот этап формирования нервной системы плода достигает своего пика на 5-6 неделе гестации. В этот период у нервной трубки появляются 3 расширенных полости (на переднем ее конце), из которых после формируются:
из 1-й (краниальной полости) - головной мозг;
из 2-й и 3-й полости - спинной мозг.
Вследствие деления на три пузыря, нервная система развивается дальше и зачаток головного мозга плода из трех пузырей превращается в пять путем деления.
Из переднего мозга образуется - конечный мозг и межуточный мозг.
Из заднего мозгового пузыря - закладка мозжечка и продолговатого мозга.
В первый триместр беременности также проходит частично нейрональная пролиферация.
Спинной мозг развивается быстрее, чем головной, и, следовательно, функционировать начинает также быстрее, отчего играет более важную роль на начальных этапах развития плода.
Но в первом триместре беременности особое внимание заслуживает процесс развития вестибулярного анализатора. Он является высокоспециализированным анализатором, который отвечает у плода за восприятие перемещения в пространстве и ощущение изменения положения. Этот анализатор формируется уже на 7 неделе внутриутробного развития (раньше других анализаторов!), а к 12-той неделе к нему уже подходят нервные волокна. Миелинизация нервных волокон начинается к моменту появления у плода первых движений - на 14 - неделе гестации. Но для проведения импульсов от вестибулярных ядер к двигательным клеткам передних рогов спинного мозга необходимо быть миелинизированным вестибуло - спинальному тракту. Его миелинизация происходит через 1-2 недели (15 - 16 неделя гестации).
Поэтому, благодаря раннему формированию вестибулярного рефлекса, при перемещении беременной женщины в пространстве плод перемещается в полости матки. Вместе с этим, перемещение плода в пространстве является «раздражающим» фактором для вестибулярного рецептора, который посылает импульсы для дальнейшего развития нервной системы плода.
Нарушения развития плода от воздействия различных факторов в этот период ведет к нарушениям вестибулярного аппарата у новорожденного ребенка.
До 2-го месяца гестации плод имеет гладкую поверхность головного мозга, покрытую эпендимным слоем, состоящим из медуллобластов. Ко 2 - му месяцу внутриутробного развития начинает формироваться кора головного мозга путем миграции нейробластов в вышележащий краевой слой, и, таким образом, формируя закладку серого вещества головного мозга.
Все неблагоприятные факторы воздействия в первый триместр развития нервной системы плода приводят к тяжелым и, в большинстве случаев, необратимым нарушениям функционирования и дальнейшего формирования нервной системы плода.
Второй триместр беременности.
Если в первом триместре беременности происходит основная закладка нервной системы, то во втором триместре происходит ее интенсивное развитие.
Нейрональная пролиферация является основным процессом онтогенеза.
На этом этапе развития возникает физиологическая водянка пузырей головного мозга. Это происходит из-за того, что спинномозговая жидкость, поступая в мозговые пузыри, расширяет их.
К концу 5-го месяца гестации образуются все основные борозды головного мозга, а также появляются отверстия Люшка, через которые спинномозговая жидкость выходит на наружную поверхность мозга и омывает его.
В течение 4 - 5 месяца развития мозга интенсивно развивается мозжечок. Он приобретает характерную ему извилистость, и делиться поперек, образуя свои основные части: переднюю, заднюю и фолликуло-нодулярные доли.
Также во втором триместре беременности проходит этап миграции клеток (5 месяц), в результате которого появляется зональность. Головной мозг плода становится более похож на головной мозг взрослого ребенка.
При воздействии неблагоприятных факторов на плод во второй период беременности, возникают нарушения, которые совместимы с жизнью, так как закладка нервной систему прошла в первом триместре. На этом этапе нарушения связанны с недоразвитием структур мозга.
Третий триместр беременности.
В этот период происходит организация и миелинизация структур головного мозга. Борозды и извилины в своем развитии подходят к завершающему этапу (7 - 8 месяц гестации).
Под этапом организации нервных структур понимают морфологическую дифференцировку и возникновение специфических нейронов. В связи с развитием цитоплазмы клеток и увеличения внутриклеточных органелл, происходит увеличение образования продуктов обмена, которые необходимы для развития нервных структур: белки, ферменты, гликолипиды, медиаторы и др. Параллельно с этими процессами протекает образование аксонов и дендритов для обеспечения синоптических контактов между нейронами.
Миелинизация нервных структур начинается с 4-5 месяца гестации и заканчивается к концу первого, началу второго года жизни ребенка, когда ребенок начинает ходить.
При воздействии неблагоприятных факторов в третьем триместре беременности, а также в течение первого года жизни, когда заканчиваются процессы миелинизации пирамидных путей, серьезных нарушений не возникает. Возможны легкие изменения структуры, которые определяются только при гистологическом исследовании.
Развитие ликвора и кровеносной системы головного и спинного мозга.
В первом триместре беременности (1 - 2 месяц гестации), когда происходит образование пяти мозговых пузырей, происходит образование сосудистых сплетений в полости первого, второго и пятого мозгового пузыря. Эти сплетения начинают секретировать высококонцентрированный ликвор, который является, по сути, питательной средой из-за большого содержания в своем составе белка и гликогена (превышает в 20 раз в отличие от взрослых). Ликвор - в этом периоде является основным источником питательных веществ для развития структур нервной системы.
Пока развитие мозговых структур поддерживает ликвор, на 3 - 4 неделе гестации образуются первые сосуды кровеносной системы, которые расположены в мягко-паутинной оболочке. Изначально содержание кислорода в артериях очень низкое, но в течение с 1 - го по 2 - й месяц внутриутробного развития кровеносная система приобретает более зрелый вид. И на втором месяце гестации кровеносные сосуды начинают врастать в мозговое вещество, образуя кровеносную сеть.
К 5 - му месяцу развития нервной системы появляются передняя, средняя и задняя мозговые артерии, которые соединены между собой анастомозами, и представляют собой завершенную структуру мозга.
Кровоснабжение спинного мозга происходит из большего количества источников, чем у головного мозга. Кровь к спинному мозгу поступает из двух позвоночных артерий, которые разветвляются на три артериальных тракта, которые, в свою очередь, идут вдоль всего спинного мозга, питая его. Передние рога получают большее количество питательных веществ.
Венозная система исключает образование коллатералей и является более изолированной, что способствует быстрому выведению конечных продуктов обмена по центральным венам на поверхность спинного мозга и выведением в венозные сплетения позвоночника.
Особенностью кровоснабжения третьего, четвертого и боковых желудочков у плода является более широкий размер капилляров, которые проходят в этих структурах. Это ведет к замедленному току крови, что способствует более интенсивному питанию.