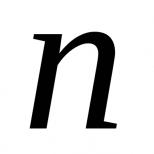Термин синапс ввел. Синапс. Классификация синапсов. Механизм передачи возбуждения в синапсе
Московский Психолого- социальный Институт (МПСИ)
Реферат по Анатомии ЦНС на тему:
СИНАПСЫ (строение, структура, функции).
Студент 1 курса Психологического факультета,
группа 21/1-01 Логачёв А.Ю.
Преподаватель:
Холодова Марина Владимировна.
2001 год.
План работы:
1.Пролог.
2.Физиология нейрона и его строение.
3.Структура и функции синапса.
4.Химический синапс.
5.Выделение медиатора.
6.Химические медиаторы и их виды.
7.Эпилог.
8.Список литературы.
ПРОЛОГ:
Наше тело - один большой часовой механизм. Он состоит из огромнейшего количества мельчайших частиц, которые расположены в строгом порядке и каждая из них выполняет определённые функции, и имеет свои неповторимые свойства. Этот механизм - тело, состоит из клеток, соединяющих их тканей и систем: все это в целом представляет собой единую цепочку, сверхсистему организма. Величайшее множество клеточных элементов не могли бы работать как единое целое, если бы в организме не существовал утонченный механизм регуляции. Особую роль в регуляции играет нервная система. Вся сложная работа нервной системы - регулирование работы внутренних органов, управление движениями, будь то простые и неосознаваемые движения (например, дыхание) или сложные, движения рук человека - все это, в сущности, основано на взаимодействии клеток между собой. Все это, в сущности, основано на передаче сигнала от одной клетке к другой. Причем, каждая клетка выполняет свою работу, а иногда имеет несколько функций. Разнообразие функций обеспечивается двумя факторами: тем, как клетки соединены между собой, и тем, как устроены эти соединения.
ФИЗИОЛОГИЯ НЕЙРОНА И ЕГО СТРОЕНИЕ:
Простейшая реакция нервной системы на внешний раздражитель - это рефлекс. Прежде всего, рассмотрим строение и физиологию структурной элементарной единицы нервной ткани животных и человека - нейрона. Функциональные и основные свойства нейрона определяются его способностью к возбуждению и самовозбуждению. Передача возбуждения осуществляется по отросткам нейрона - аксонам и дендритам.
Аксоны - более длинные и широкие отростки. Они обладают рядом специфических свойств: изолированным проведением возбуждения и двусторонней проводимостью.
Нервные клетки способны не только воспринимать и перерабатывать внешнее возбуждение, но и самопроизвольно выдавать импульсы, не вызванные внешним раздражением (самовозбуждение). В ответ на раздражение, нейрон отвечает импульсом активности - потенциалом действия, частота генерации которых колеблется от 50-60 импульсов в секунду (для мотонейронов), до 600-800 импульсов в секунду (для вставочных нейронов головного мозга). Аксон заканчивается множеством тоненьких веточек, которые называются терминалями. С терминалей импульс переходит на другие клетки, непосредственно на их тела или чаще на их отростки дендриты. Количество терминалей у аксона, может достигать до одной тысячи, которые оканчиваются в разных клетках. С другой стороны, типичный нейрон позвоночного имеет от 1000 до 10000 терминалей от других клеток.
Дендриты - более короткие и многочисленные отростки нейронов. Они воспринимают возбуждение от соседних нейронов и проводят его к телу клетки. Различают мякотные и безмякотные нервные клетки и волокна.
Мякотные волокна - входят в состав чувствительных и двигательных нервов скелетной мускулатуры и органов чувств Они покрыты липидной миелиновой оболочкой. Мякотные волокна более «быстродействующие»: в таких волокнах диаметром 1-3,5 микромиллиметра, возбуждение распространяется со скоростью 3-18 м/с. Это объясняется тем, что проведение импульсов по миелинизированному нерву происходит скачкообразно. При этом потенциал действия «перескакивает» через участок нерва, покрытый миелином и в месте перехвата Ранвье (оголенный участок нерва), переходит на оболочку осевого цилиндра нервного волокна. Миелиновая оболочка является хорошим изолятором и исключает передачу возбуждения на соединение, параллельно идущие нервные волокна.
Безмякотные волокна - составляют основную часть симпатических нервов. Они не имеют миелиновой оболочки и отделены друг от друга клетками нейроглии.
В безмякотных волокнах роль изоляторов выполняют клетки нейроглии (нервной опорной ткани). Швановские клетки - одна из разновидностей глиальных клеток. Помимо внутренних нейронов, воспринимающих и преобразующих импульсы, поступающие от других нейронов, существуют нейроны, воспринимающие воздействия непосредственно из окружающей среды - это рецепторы, а так же нейроны, непосредственно воздействующие на исполнительные органы - эффекторы, например, на мышцы или железы. Если нейрон воздействует на мышцу, его называют моторным нейроном или мотонейроном. Среди нейрорецепторов различают 5 типов клеток, в зависимости от вида возбудителя:
- фоторецепторы, которые возбуждаются под воздействием света и обеспечивают работу органов зрения,
- механорецепторы, те рецепторы, которые реагируют на механические воздействия. Они располагаются в органах слуха, равновесия. Осязательные клетки также являются механорецепторами. Некоторые механорецепторы располагаются в мышцах и измеряют степень их растяжения.
- хеморецепторы - избирательно реагируют на присутствие или изменение концентрации различных химических веществ, на них основана работа органов обоняния и вкуса,
- терморецепторы, реагируют на изменение температуры либо на ее уровень - холодовые и тепловые рецепторы,
- электрорецепторы реагируют на токовые импульсы, и имеются у некоторых рыб, амфибий и млекопитающих, например, у утконоса.
Исходя из выше сказанного, хотелось бы отметить, что долгое время среди биологов, изучавших нервную систему, существовало мнение, что нервные клетки образуют длинные сложные сети, непрерывно переходящие одна в другую.
Однако в 1875 году, итальянский ученый, профессор гистологии университета в Павии, придумал новый способ окраски клеток - серебрение. При серебрении одной из тысяч лежащих рядом клеток окрашивается только она - единственная, но зато полностью, со всеми своими отростками. Метод Гольджи сильно помог изучению строения нервных клеток. Его использование показало, что, не смотря на то, что клетки в головном мозгу расположены чрезвычайно близко друг к другу, и их отростки перепутаны, все же каждая клетка четко отделяется. То есть мозг, как и другие ткани, состоит из отдельных, не объединенных в общую сеть клеток. Этот вывод был сделан испанским гистологом С. Рамон-и-Кахалем, который тем самым распространил клеточную теорию на нервную систему. Отказ от представления об объединенной сети, означал, что в нервной системе импульс переходит с клетки на клетку не через прямой электрический контакт, а через разрыв.
Когда в биологии стал использоваться электронный микроскоп, который был изобретен в 1931 году М. Кноллем и Э. Руска, эти представления о наличии разрыва получили прямое подтверждение.
СТРУКТУРА И ФУНКЦИИ СИНАПСА:
Каждый многоклеточный организм, каждая ткань, состоящая из клеток, нуждается в механизмах, обеспечивающих межклеточные взаимодействия. Рассмотрим, как осуществляются межнейронные взаимодействия. По нервной клетке информация распространяется в виде потенциалов действия. Передача возбуждения с аксонных терминалей на иннервируемый орган или другую нервную клетку происходит через межклеточные структурные образования - синапы (от греч. «Synapsis» -соединение, связь). Понятие синапс было введено английским физиологом Ч. Шеррингтоном в 1897 году, для обозначения функционального контакта между нейронами. Следует отметить, что еще в 60-х годах прошлого столетия И.М. Сеченов подчеркивал, что вне межклеточной связи нельзя объяснить способы происхождения даже самого нервного элементарного процесса. Чем сложнее устроена нервная система, и чем больше число составляющих нервных мозговых элементов, тем важнее становится значение синаптических контактов.
Различные синаптические контакты отличаются друг от друга. Однако при всем многообразии синапсов существуют определенные общие свойства их структуры и функции. Поэтому сначала опишем общие принципы их функционирования.
Синапс - представляет собой сложное структурное образование, состоящее из пресинаптической мембраны (чаще всего это концевое разветвление аксона), постсинаптической мембраны (чаще всего это участок мембраны тела или дендрита другого нейрона), а так же синаптической щели.
Механизм передачи через синапс долгое время оставался невыясненным, хотя было очевидно, что передача сигналов в синаптической области резко отличается от процесса проведения потенциала действия по аксону. Однако в начале XX века была сформулирована гипотеза, что синаптическая передача осуществляется или электрическим или химическим путем. Электрическая теория синаптической передачи в ЦНС пользовалась признанием до начала 50-х годов, однако она значительно сдала свои позиции после того, как химический синапс был продемонстрирован в ряде периферических синапсов. Так, например, А.В. Кибяков, проведя опыт на нервном ганглии, а также использование микроэлектродной техники для внутриклеточной регистрации синаптических потенциалов
нейронов ЦНС позволили сделать вывод о химической природе передачи в межнейрональных синапсах спинного мозга.
Микроэлектродные исследования последних лет показали, что в определенных межнейронных синапсах существует электрический механизм передачи. В настоящее время стало очевидным, что есть синапсы, как с химическим механизмом передачи, так и с электрическим. Более того, в некоторых синаптических структурах вместе функционируют и электрический и химический механизмы передачи - это так называемые смешанные синапсы.
Межнейронные контакты - синапсы: строение, функция и эволюция .
Санкт-Петербургский государственный университет. Санкт-Петербург, Центр «Интеллект» Лисий нос. ЛО.
Биология в школе 2016, № 7, с. 3-12.
Аннотация
В статье приведены современные сведения о структуре, принципах работы и эволюции межнейронных контактов (синапсов) различного вида. Подробно разбирается морфология синапсов. Особое внимание уделено вопросам молекулярных механизмов работы синапсов и их эволюции в процессе становления и развития нервной системы у животных.
Ключевые слова:
нервная система, нейрон, синапс, медиаторы, синаптические рецепторы
Введение
Долгое время в нейробиологии боролись два представления о принципах клеточной организации нервной системы. По одному из них постулировалось, что в нервной системе возможны прямые контакты (типа синцития) между отростками нервных клеток. Приверженцем этой теории был знаменитый итальянский нейрогистолог Камилло Гольджи (1843– 1926). Вторая точка зрения, поддержанная выдающимся испанским нейрогистологом Сантьяго Рамон-и-Кахалем (1852-1934) постулировала, что между нервными клетками существует система специализированных контактов – синапсов. Интересно, что оба они одновременно были удостоены Нобелевской премии (1906) за исследования нервной системы. Только с появлением современных электронных микроскопов вторая точка зрения получила окончательное подтверждение. Сам термин - синапс (греч. sinapsis – соединение, связь ) был введен известным английским нейрофизиологом Ч. Шериннгтоном еще в конце 19-го века.
Таким образом, в конце ХХ – века окончательно сложилась «нейронная теория строения нервной системы », основными положениями которой стали тезисы о том, что нейрон – это элементарная единица нервной ткани и что нейроны соединяются друг с другом или с другими клетками организма посредством специальных межнейронных прерывистых контактов – синапсов .
Классификация синапсов.
Классифицировать синапсы можно исходя из разных их параметров и свойств.
По способу передачи межнейронные контакты можно разделить на: химические, электрические и смешанные (электро-химические).
- По той части нейрона, которая образует контакт, выделяют наиболее часто встречающиеся аксо-дендритные или аксо-шипиковые контакты (шипики – многочисленные микроскопические выросты на дендритах). Реже бывают аксо-аксональные, аксо-соматические и дендро-дендритные синапсы . Последняя группа синапсов имеет значение для обеспечения процессов регуляции деятельности нейронов (например: при взаимодействии аксонных терминалей с аксоном собственного нейрона) или для осуществления тормозных взаимодействий в нейронных цепях (например, при пресинаптическом торможении). В принципе любая часть нейрона может образовать контакт с любой частью другого нейрона. Нейроны также могут образовывать синапсы с клетками других тканей и органов (например: нейро-мышечные контакты). Связи между нейронами могут осуществляться как одиночными синапсами различного вида, так и сложными комплексами типа "гломерул". Синаптические комплексы построены по двум принципам: конвергенции и дивергенции . В центре конвергентного синаптического комплекса находится один постсинаптический элемент – тело или отросток нейрона, на котором оканчивается несколько пресинапсов различного происхождения и типа. В дивергентном комплексе отношения обратные: одна пресинаптическая часть контакта – аксон, оказывает влияние на несколько постсинаптических элементов.
По механизму действия на соседние нейроны синапсы разделяются на возбуждающие или тормозные (см. ниже ).
Химические синапсы (строение, принцип работы, свойства). Этот вид межнейронных контактов широко представлен в нервной системе всех животных – от низших беспозвоночных до высших млекопитающих и человека. У высших животных (как позвоночных, так и беспозвоночных) они преобладают в нервной системе.
Принципиальная схема строения химического синапса едина: та часть нервной клетки (аксон, дендрит или участок тела нейрона), которая образует контакт, называется пресинапсом , затем идет синаптическая щель (шириной от 10 до 500 нм), а та часть, на которой оканчивается контакт – постсинапсом . (Рис. 1).
Рис.1 Электронная фотография химического синапса.
Обозначения: 1 – пресинапс, 2 – постсинапс, 3 – скопления синаптических пузырьков, 4 – пресинаптическое утолщение (активная зона синапса), 5 – постсинаптическое утолщение, 6 –митохондрия в пресинапсе. Масштаб 200 нм.
Основной принцип работы химического синапса – использование специальных химических веществ (медиаторов, трансмиттеров) для передачи сигнала от одного нейрона другому. Медиаторы находятся в пресинапсе в специальных мембранных структурах – синаптических пузырьках . Показано, что популяция синаптических пузырьков неоднородна. Большая часть их образует т. н. «резервный пул», который располагается в отдалении от пресинаптической мембраны и «рабочий пул», который используется в работе синапса в первую очередь. Пузырьки претерпевают в процессе работы синапса определенный цикл, по мере которого они используются в работе синапса, а затем вновь заполняются медиатором/ми. Также в пресинапсе находятся элементы цитоскелета и транспортной системы клетки (микротрубочки, актиновые филаменты, миозин), митохондрии, ферментные системы для синтеза медиатора. (Рис. 2).

Рис. 2 Схема строения химического синапса (аксо-шипиковый синапс).
Обозначения: 1 – миелиновая оболочка аксона; 2 – аксон; 3 – аксонная терминаль (пресинапс); 4 транспорт синаптических пузырьков по аксону или в самой терминали; 5 – синаптический пузырек с медиатором/ми; 6 – движение пузырька к пресинапстической мембране; 7 - Са+2 каналы (активированные в момент прихода импульса); 8 – слияние (fusion) пузырька с пресинаптической мембраной в активной зоне синапса и экзоцитоз медиатора в синаптическую щель; 9 – диффузия медиатора в синаптической щели к постсинаптической мембране; 10 – пресинаптические рецепторы (в том числе к собственному медиатору/ам); 11 – эндоцитоз пузырьков; 12,13 – рециклинг пузырьков в пресинапсе с образованием эндосомы и новых пузырьков.
Известно несколько десятков химических веществ, выполняющих функции нейромедиаторов или нейромодуляторов. Это могут быть: аминокислоты (глутамат, глицин, ГАМК, таурин и др.); амины (ацетилхолин, гистамин, серотонин, допамин, адреналин , норадренаил и др.); белки (энкефалины, эндорфины, вещество Р, VIP - нейропептид, нейротензин, ряд других нейрогормонов); пуриновые соединения (АТФ, ГТФ, аденозин, инозин) и даже газообразные вещества (NO, CO, H2S).
Газообразные посредники имеют ряд свойств, отличающих их от классических медиаторов. Все они легко проникают через мембрану, выделяются из любого участка клетки, не запасаются в синаптических пузырьках и не освобождаются экзоцитозом и являются коротко живущими. Клеточные эффекты газов опосредуются либо через систему внутриклеточных посредников, либо через прямое влияние на субъединицы ионных каналов, белки экзоцитоза, внутриклеточные ферменты. В роли нейромедиаторов и нейромодуляторов газы имеют преимущества перед другими посредниками по скорости синтеза и выделения, степени проницаемости через мембрану и широкому спектру мишеней. Особенности действия газов позволяют предполагать их важную роль в процессах развития нервной системы, формировании кратковременных и долговременных изменений в синаптических структурах, связанных с процессами памяти и обучения.
При этом нейроны могут синтезировать и выделять в своих окончаниях целый набор медиаторов и комедиаторов (например, в ацетилхолиновых синапсах в качестве комедиаторов могут присутствовать: энкефалин, VIP, вещество P, соматостатин или нейротензин).
Вещества, которые претендуют на роль медиатора, должны соответствовать нескольким критериям:
Они должны синтезироваться нейроном и храниться в синапсах;
При поступлении нервного импульса выделяться в синаптическую щель и избирательно связываться со специфическими рецепторами на постсинаптической мембране другого нейрона;
Вызывать соответствующую физиологическую реакцию;
При введении в нервную систему извне оказывать такой же физиологический эффект, как и эндогенные медиаторы.
Синтез медиаторов идет в теле клетки, а затем с помощью транспортной системы нейрона пузырьки, заполненные медиатором (или пустые), поставляются по аксону в пресинапс. Часть медиатора синтезируется непосредственно в синапсе и заполняет синаптические пузырьки. Поскольку в одном синапсе может быть несколько медиаторов, то они могут находиться как в отдельных синаптических пузырьках, так и в одном (пузырьковая фракция). Кроме того часть медиатора/ов обнаруживается непосредственно в цитоплазме пресинапса (цитоплазматическая фракция). Обе эти фракции могут обмениваться друг с другом и участвовать в работе синапса. Количество медиатора, находящегося в одном пузырьке, получило наименование «квант». Например, в пузырьке ацетилхолинового синапса содержится примерно 10000 молекул ацетилхолина. Выброс медиатора с синаптическую щель происходит квантами, но для возникновения нервного импульса необходим выброс одновременно множества квантов.
В нервной системе позвоночных и беспозвоночных животных используются одни и те же медиаторы . Различия касаются только набора используемых медиаторов. В нервной системе низших животных встречается меньшее разнообразие медиаторов, чем в нервной системе высших позвоночных и беспозвоночных животных. Большее разнообразие дает возможность комбинировать наборы медиаторов в синапсах и создавать более сложные нейронные сети.
Важно отметить, что термины "нейромедиатор", "нейромодулятор", "нейрогормон" отражают скорее механизм взаимодействия этих соединений с клетками-мишенями, чем их химическую природу. Одно и то же вещество может выступать как в роли медиатора, так и нейрогормона. Многие нейропептиды, например энкефалины и эндорфины, ранее рассматриваемые только как нейрогормоны, выделяются аксонными терминалями и выступают в роли нейромедиаторов. Другие действуют не только через межнейронные сипапсы, но и выделяются нейросекреторными и эндокринными клетками, выступая как типичные гормоны (адреналин, дофамин, серотонин и др.)
Важнейшей частью пресинапса является пресинаптическая мембрана с ее специализированными участками, названными активными зонами синапса , где происходит контакт синаптичеких пузырьков с мембраной и экзоцитоз медиатора в синаптическую щель. Активная зона не занимает всю площадь синаптического контакта и может меняться в зависимости от активности работы нейрона.
В состав активной зоны синапса - помимо самой пресинаптической мембраны, входят т. н. «пресинаптические субмембранные утолщения » имеющие в проекции гексагональное расположение и отличающиеся чрезвычайно сложным устройством. В их состав входит около 100 белков, наиболее важные из которых можно объединить в три комплекса. Первый комплекс предназначен для формирования основы цитоматрикса активной зоны. Второй белковый комплекс взаимодействует с мембраной пресинапса и регулирует экзоцитоз синаптических пузырьков. Третий, ключевой в этой белковой системе т. н. «SNARE – комплекс», обеспечивает подход (docking) и слияние (fusion) синаптических пузырьков с пресинаптической мембраной. Он включает такие важные синаптические белки как: синтаксин, синапсин, синаптобревин (VAMP) и белок SNARE-25. Важность этого комплекса подчеркивается фактом полного нарушения работы синапса при взаимодействии белков SNARE – комплекса с рядом нейротоксинов (ботулином или столбнячным токсином).
Со стороны пре - и постсинаптической мембраны в синаптическую щель выступают молекулы межклеточной адгезии (катенины, кадгерины, нейрексины, нейролигины и др.). Их роль состоит в связывании и закреплении пре - и постсинаптических мембран. Кроме того, в синаптической щели могут находиться ферменты, предназначенные для дезактивации медиатора/ров после выполнения ими своей функции.
Постсинапс является важнейшей частью синаптического контакта и включает несколько компонентов: собственно постсинаптическую мембрану со встроенными в нее постсинаптическими рецепторами и ионными каналами, субмембранный комплекс (постсинатическое утолщение ) и элементы цитоскелета постсинапса. Вся эта структура предназначена для стабилизации постсинаптических рецепторов и ионных каналов в мембране и формирования нового нервного импульса (постсинатическое возбуждение или торможение ).
Важно отметить, что знак работы синапса не зависит от химической природы медиатора, а связан со свойствами постсинаптических рецепторов. Этот важнейший факт был установлен автралийским ученым, лауреатом Нобелевской премии 1963 года.
Постсинаптические рецепторы являются сложными белковыми комплексами, встроенными в постсинаптическую мембрану. Выделяют три основных вида таких рецепторов: ионотропные, метаботроные и каталитические.
Ионотропные рецепторы представлены мембранными белковыми комплексами, состоящими, как правило, из 4-5 белковых субъединиц с ионным каналом в центре. При связывании медиатора с таким рецептором одновременно открывается ионный канал, происходит перераспределение ионных потоков внутри и снаружи мембраны, вследствие чего изменяется трансмембранный потенциал и возникает нервный импульс (постсинатический потенциал действия). (Рис 3, а).

Рис. 3 Схема строения ионотропного (А) и метаботропного (В) синаптических рецепторов, Б – субъединицы ионотропного никотинового ацетилхолинового (N-Ah) рецептора (объяснения в тексте).
Ионотропные рецепторы открываются почти мгновенно (время реакции ~10 мкс), но остаются открытыми лишь в течение нескольких миллисекунд. Ионотропные рецепторы классифицируются по типу медиатора , с которым они связываются, и по типу ионного канала . Если рецептор работанет на Na+ или Ca+2 - канале, то при их активации , происходит вход ионов внутрь постсинапса, развивается деполяризация мембраны и возникает возбуждающий постсинаптический потенциал (ВПСП). Если рецептор работает на хлорном (Cl- ) канале, то происходит гиперполяризация мембраны и возникает тормозный постсинаптический потенциал (ТПСП), препятствующий возникновению потенциала действия . (ПД). Синаптическое торможение играет важную физиологическую роль в ЦНС, ограничивая избыточное возбуждение в нейронных сетях.
Одним из наиболее изученных синаптических рецепторов является ионотропный никотиновый ацетилхолиновый рецептор ( n AhR) , работающий в нервно-мышечном синапсе. Он представляет собой мембранный белковый комплекс, состоящий из 5-ти интегральных белков и ионного К+/Na+ - канала. В его состав входят: две α 1 - и по одной: β , γ, δ - субъединицы. (Рис. 3,б) Медиатор (две молекулы ацетилхолина) соединяются с двумя α 1 – субъединицами рецептора.
В нервной системе существует несколько модификаций ацетилхолинового рецептора, различающихся по типу и соотношению субъединиц, входящих в состав рецептора. Например, в ЦНС существует ацетилхолиновый рецептор, состоящий из пяти α-субъединиц 7-типа . (α7)5. Важно отметить, что разные модификации ацетилхолинового ионотропного рецептора встречаются в разных отделах нервной системы и обладают разными функциональными характеристиками. Таким образом, создается возможность комбинировать набор рецепторов в разных синапсах и нейронах.
Метаботропные рецепторы представляют собой сложный молекулярный комплекс, где рецепторная часть и ионный канал пространственно разобщены в мембране постсинапса (Рис. 3,в ). В связи с этим необходим «посредник/и, мессенджер » для связи их друг с другом и дальнейшей передачи сигнала на мембрану или вглубь клетки.
После взаимодействия медиатора с рецептором происходит активация т. н. G – белка . G - белок обладает ферментативной активностью и активирует мембранный белок аденилатциклазу, которая, в свою очередь, превращает АТФ в молекулы циклического аденозинмонофосфата (цАМФ - вторичный посредник ) . Одна молекула аденилатциклазы вызывает образование множества молекул цАМФ. Молекулы цАМФ активируют цАМФ-зависимую протеинкиназу, которая фосфорилирует белки ионного канала и он открывается.
Эффект всех метаботропных рецепторов опосредуется через те или иные системы вторичных посредников. В качестве вторичных посредников могут выступать многие соединения: циклический аденозин - или гуанинмонофосфат (цАМФ, цГМФ); инозитол-3-фосфат (ИФ3); диацилглицерид; тирозин; ионы Са+2 и др. Помимо мембранных протеинкиназ они могут активировать соответствующие внутриклеточные протеинкиназы, которые путем фосфорилирования цитоплазматических или ядерных белков запускают разнообразные внутриклеточные реакции. Так, в частности, действуют многие гормоны.
Метаботропные рецепторы сохраняют активированное состояние в течение секунд или минут после связывания с медиатором. Поэтому они имеют более длительные эффекты, чем ионотропные рецепторы.
Как и ионотропный никотиновый рецептор (nAhR), метаботропный мускариновый ацетилхолиновый рецептор (mAhR) имеет несколько разновидностей, различающихся по вторичным посредникам и типам ионных каналов (Табл. 1).
Табл. 1 . Некоторые характеристики метаботропных ацетилхолиновых рецепторов разного вида (М1-М5) и их распределение в организме человека. Количество плюсов отражает степень встречаемости рецептора в данной структуре/органе.
Тип G - белка | |||||
Вторичный посредник | |||||
Ионный канал | |||||
Центральная нервная система |
|||||
Неокортекс | |||||
Мозжечок | |||||
Гиппокамп | |||||
Гипоталамус | |||||
Спинной мозг | |||||
Периферическая нервная система и органы |
|||||
Гладкая мускулатура | |||||
Нейроны симпатических ганглиев |
Также как и никотиновые (nAhR), мускариновые (mAhR) рецепторы разных подтипов могут располагаться как на разных нервных клетках, так и на одном нейроне. Например, на пирамидном нейроне гиппокампа найдены все пять подтипов mAhR. (см. Таб. 1 ).
Другие медиаторы, встречающиеся в нервной системе животных и человека, имеют свои разновидности ионотропных и метаботропных рецепторов. Порой весьма многочисленные.
Каталитические рецепторы , также распространенные в нервной системе позвоночных и беспозвоночных животных, представляют собой белковые структуры, способные, с одной стороны, связываться с медиатором, с другой стороны, обладают ферментативной активностью. Большая часть таких рецепторов представлена тирозинкиназами, которые способны при активации фосфорилировать как сам белок–рецептор (автофосфорилирование), так и белки в цитоплазме, запуская каскад биохимических реакций в клетке.
Исходя из особенностей строения и принципов работы, химические синапсы обладают рядом общих свойств . Они односторонние (синаптические пузырьки находятся только в пресинапсе); медленные (процессы экзоцитоза медиатора, его взаимодействия с рецепторами и т. д. занимают много времени); утомляемые (развитие утомления связано с истощением запасов нейромедиатора, который может израсходоваться в несколько минут, а иногда и секунд).
Итак, основные этапы передачи в химическом синапсе можно кратко описать следующим образом:
1. Нервный импульс, идущий по мембране аксона, достигает пресинаптической терминали.
2. Деполяризация мембраны аксонного окончания приводит к активации расположенных на ней потенциалзависимых Са+2 - каналов и ионы Са+2 устремляются внутрь синапса, активируя систему транспорта синаптических пузырьков к активной зоне пресинаптической мембраны
3. Синаптические пузырьки взаимодействуют с белками активной зоны синапса и путем экзоцитоза выделяют медиатор в синаптическую щель, где они диффундируют к постсинаптической мембране.
4. Медиатор взаимодействует с рецепторами постсинаптической мембраны.
5. Активация ионных каналов приводит к де - или гиперполяризации постсинаптической мембраны, в результате чего формируется возбуждающий (ВПСП) или тормозный (ТПСП) постсинаптический потенциал.
6. После прекращения взаимодействия медиатора с рецептором происходит процесс его инактивации путем обратного захвата медиатора пресинапсом, либо расщеплением его ферментами, находящимися в синаптической щели, либо захватом медиатора и его компонентов глиальными клетками.
Электротонические (электрические) синапсы . Впервые явление прямой электрической передачи импульса в синапсах было продемонстрировано при изучении гигантских нервных волокон ракообразных в 1959 году. В настоящее время показано, что электрические синапсы представляют собой одну из разновидностей межклеточных контактов – т. н. щелевых контактов (gap j u nction). (Рис. 4.)

Рис.4 . Электрический синапс.
Электронная фотография дендро-дендриного контакта (А), схема строения эл. синапса (Б); строение коннексонов (В) (объяснения в тексте).
Основу его составляют сложные молекулярные комплексы – коннексоны , каждый из которых образован своеобразной «розеткой» из шести белков – коннексинов (connexin - Cx ). В центре такого комплекса находится канал, который может пропускать ионы, чем и объясняются электрические свойства контакта. Коннексоны двух соседних мембран контактируют в узкой синаптической щели (2-3 нм), разделяющей соседние нейроны. Важно подчеркнуть, что никакого слияния мембран соседних клеток при этом не происходит.
Коннексины, образующие коннексоны в электрических синапсах у разных типах нейронов и глиальных клеток, различаются по молекулярному весу и кодируются разными генами. Так, например, коннексоны, образующие контакты между астроцитами (разновидность глиальных клеток) состоят из коннексинов - Cx43, а между интернейронами коры больших полушарий – Cx36. Также показано, что в состав коннексонов могут входить разные комбинации коннексинов. Естественно, что это определяет различия в функциональных параметрах данных контактов, например, разную чувствительность к гипер - или деполяризации мембран контакта или разную проницаемость для анионов или катионов.
Такая структура электрических синапсов определяет и основные функциональные особенности данных контактов, существенно отличающих их от химических синапсов: возможность проводить импульс в обоих направлениях, большая скорость проведения импульса, неутомляемость.
Электрические синапсы распространены и в нервной системе беспозвоночных животных и имеют сходное строение с таковыми у позвоночных. Белки, образующие коннексоны у беспозвоночных животных получили наименование иннексины ( innexsin – Inx ) и паннексины ( pannexin – Panx ). Они во многом гомологичны коннексинам позвоночных животных.
Функциональное предназначение электрических синапсов в нервной системе состоит в возможности быстрой координации и согласованной деятельности групп нейронов в тех или иных нервных центрах. Особенно их много в ганглиях нервной системы беспозвоночных и в ряде отделов нервной системы позвоночных животных, чья деятельность связана с осуществлением и регуляцией быстрых врожденных рефлексов (например: в центрах спинного мозга и ствола мозга). В связи с этим, одни исследователи рассматривают электротонические синапсы как первичные в эволюции нервной системы и указывают на существование в филогенезе процесса замещения электротонических контактов химическими. Другие считают, что как электрические, так и химические синапсы возникли в эволюции нервной системы очень рано и имеют широкое распространение в нервных центрах животных всех филогенетических групп. Преобладание же в ряде нервных центров того или иного типа синаптической передачи определяется функциональными задачами данного центра, а не его филогенетическим происхождением. Эта точка зрения базируется на фактах обнаружения типичных химических и электрических синапсов в нервной системе низших многоклеточных животных с одной стороны, и в высших интегративных центрах конечного мозга млекопитающих и птиц с другой. Более того, показано, что синапсы, образуемые одним аксоном, могут работать с использованием и химического и электротонического механизмов (т. н. смешанные электро-химические синапсы ). Интересно, что в смешанных синапсах одновременно могут работать как химический, так и электрический компоненты контакта.
Помимо участия в проведении нервного импульса электротонические контакты выполняют и другую важную функцию в нервной системе. Размер поры в коннексонах позволяет проходить через них не только ионам, но и ряду молекул весом до 1kD. Среди них могут быть такие важные молекулы как: вторичные посредники (цАМФ, цГМФ, Са2+, ИФ3), витамины , ганглиозиды, простогландины, некоторые медиаторы и даже низкомолекулярные РНК. Это явление получило название «транссинаптический перенос » и играет важную роль в метаболизме нейронов. Электрическая передача довольно устойчива (по сравнению с химическими синапсами) к различным воздействиям на нервную систему (фармакологические влияния, кислородное голодание, понижение температуры, ионный дисбаланс).Таким образом, видно, что электрические синапсы являются равноправными элементами межнейронных взаимодействий и необходимы для нормального функционирования нервной системы всех животных и человека.
Литература
1. , Каменская нервной системы. Уч. пособие. – Л.: Изд-во ЛГУ, 1985. – 160 с.
3. , Гришин и синапсы. Уч. пособие. – Казань.: Изд-во КГУ, 2003. – 130 с.
4. , Кириленкова и ткани. Уч. пособие. – М.: Дрофа, 2008. – 288 с.
5. Руководство по гистологии (под ред.) – 2-е изд. Т.1 – СПб.: Спецлит 2011. – 831 с.
6. Structural and functional organization of the synapse (J. W Hell, M. D Ehlers, eds.) – Springer Publ., N-Y-Berlin, 2008.
Академик Российской и Европейской академии естествознания, доктор биологических наук, профессор Санкт-Петербургского государственного университета, учитель биологии бысшей категории; преподаватель ГБОУ ДОД «Центр Интеллект», Ленинградская область , пос. Лисий нос.
Мышечную и железистую клетку передается посредством специального структурного образования — синапса.
Синапс — структура, обеспечивающая проведение сигнала от одной к другой. Термин был введен английским физиологом Ч. Шеррингтоном в 1897 г.
Строение синапса
Синапсы состоят из трех основных элементов: пресинаптической мембраны, постсинаптической мембраны и синаптической щели (рис. 1).
Рис. 1. Строение синапса: 1 — микротрубочки; 2 — митохондрии; 3 — синаптические пузырьки с медиатором; 4 — пресинаптическая мембрана; 5 — постсинаптическая мембрана; 6 — рецепторы; 7 -синаптическая щель
Некоторые элементы синапсов могут иметь и другие названия. Например, синаптическая бляшка — это синапс между , концевая пластинка — постсинаптическая мембрана , моторная бляшка — пресинаптическое окончание аксона на мышечном волокне.

Пресинаптическая мембрана покрывает расширенное нервное окончание, которое представляет собой нейросекреторный аппарат. В пресинаптической части находятся пузырьки и митохондрии, обеспечивающие синтез медиатора. Медиаторы депонируются в гранулах (пузырьках).
Постсинаптическая мембрана - утолщенная часть мембраны клетки, с которой контактирует пресинаптическая мембрана. Она имеет ионные каналы и способна к генерации потенциала действия. Кроме того, на ней расположены специальные белковые структуры — рецепторы, воспринимающие действие медиаторов.
Синаптическая щель представляет собой пространство между пресинаптической и постсинаптической мембранами, заполненное жидкостью, близкой по составу к .

Рис. Строение синапса и процессы, осуществляемые в ходе синаптической передачи сигнала
Виды синапсов
Синапсы классифицируются по местоположению, характеру действия, способу передачи сигнала.
По месту положения выделяют нервно-мышечные синапсы, нервно-железистые и нейро-нейрональные; последние, в свою очередь, делятся на аксо-аксональные, аксо-дендритические, аксо-соматические, дендро-соматические, дендро-дендротические.
По характеру действия на воспринимающую структуру синапсы могут быть возбуждающими и тормозящими.
По способу передачи сигнала синапсы делятся на электрические, химические, смешанные.
Таблица 1. Классификация и виды синапсов

Классификация синапсов и механизм передачи возбуждения
Синапсы классифицируют следующим образом:
- по местоположению — периферические и центральные;
- по характеру их действия — возбуждающие и тормозящие;
- по способу передачи сигналов — химические, электрические, смешанные;
- по медиатору, с помощью которого осуществляется передача, — холинергические, адренергические, серотонинергические и т.д.
В возбуждение передается с помощью медиаторов (посредников).
Медиаторы — молекулы химических веществ, которые обеспечивают передачу возбуждения в синапсах. Другими словами химические вещества, участвующие в передаче возбуждения или торможения от одной возбудимой клетки к другой.
Свойства медиаторов
- Синтезируются в нейроне
- Накапливаются в окончании клетки
- Выделяются при появлении иона Са2+ в пресинаптическом окончании
- Оказывают специфическое действие на постсинаптическую мембрану
По химическому строению медиаторы можно подразделить на амины (норадреналин, дофамин, серотонин), аминокислоты (глицин, гамма-аминомасляная кислота) и полипептиды (эндорфины, энкефалины). Ацетилхолин известен в основном как возбуждающий медиатор и содержится в различных отделах ЦНС. Медиатор находится в пузырьках пресинаптического утолщения (синаптической бляшки). Медиатор синтезируется в клетках нейрона и может ресинтезироваться из метаболитов его расщепления в синаптической щели.
При возбуждении терминалей аксона происходит деполяризация мембраны синаптической бляшки, вызывающая поступление ионов кальция из внеклеточной среды внутрь нервного окончания через кальциевые каналы. Ионы кальция стимулируют перемещение синаптических пузырьков к пресинаптической мембране, их слияние с ней и последующий выход медиатора в синаптическую щель. После проникновения в щель медиатор диффундирует к постсинаптической мембране, содержащей на своей поверхности рецепторы. Взаимодействие медиатора с рецепторами вызывает открытие натриевых каналов, что способствует деполяризации постсинаптической мембраны и возникновению возбуждающего постсинаптического потенциала. В нервно-мышечном синапсе этот потенциал называется потенциалом концевой пластинки. Между деполяризованной постсинаптической мембраной и соседними с ней поляризованными участками этой же мембраны возникают местные токи, которые деполяризуют мембрану до критического уровня с последующей генерацией потенциала действия. Потенциал действия распространяется по всем мембранам, например, мышечного волокна и вызывает его сокращение.
Выделившийся в синаптическую щель медиатор связывается с рецепторами постсинаптической мембраны и подвергается расщеплению соответствующим ферментом. Так, холинэстераза разрушает медиатор ацетилхолин. После этого некоторое количество продуктов расщепления медиатора поступает в синаптическую бляшку, где из них снова ресинтезируется ацетилхолин.
В организме имеются не только возбуждающие, но и тормозные синапсы. По механизму передачи возбуждения они сходны с синапсами возбуждающего действия. В тормозных синапсах медиатор (например, гамма-аминомасляная кислота) связывается с рецепторами постсинаптической мембраны и способствует открытию в ней . При этом активизируется проникновение этих ионов внутрь клетки и развивается гиперполяризация постсинаптической мембраны, обусловливающая возникновение тормозного постсинаптического потенциала.
В настоящее время выяснено, что один медиатор может связываться с несколькими различными рецепторами и индуцировать различные реакции.
Химические синапсы
Физиологические свойства химических синапсов
Синапсы с химической передачей возбуждения обладают определенными свойствами:
- возбуждение проводится в одном направлении, так как медиатор выделяется только из синаптической бляшки и взаимодействует с рецепторами на постсинаптической мембраны;
- распространение возбуждения через синапсы происходит медленнее, чем по нервному волокну (синаптическая задержка);
- передача возбуждения осуществляется с помощью специфических медиаторов;
- в синапсах изменяется ритм возбуждения;
- синапсы способны утомляться;
- синапсы обладают высокой чувствительностью к различным химическим веществам и гипоксии.
Одностороннее проведение сигнала. Сигнал передается только от пресинаптической мембраны к постсинаптической. Это вытекает из особенностей строения и свойств синаптических структур.
Замедленная передача сигнала. Обусловлена синаптической задержкой в передаче сигнала с одной клетки на другую. Задержка вызывается временными затратами на процессы выброса медиатора, его диффузии к постсинаптической мембране, связывания с рецепторами постсинаптической мембраны, деполяризации и преобразования постсинаптического потенциала в ПД (потенциал действия). Длительность синаптической задержки колеблется от 0,5 до 2 мс.
Способность к суммации эффекта от приходящих к синапсу сигналов. Такая суммация проявляется, если последующий сигнал приходит к синапсу через короткое время (1- 10 мс) после предыдущего. В таких случаях амплитуда ВПСП возрастает и на постсинаптическом нейроне может генерироваться большая частота ПД.
Трансформация ритма возбуждении. Частота нервных импульсов, приходящих к пресинаптической мембране, обычно не соответствует частоте ПД, генерируемых постсинаптическим нейроном. Исключение составляют синапсы, передающие возбуждение с нервного волокна на скелетную мышцу.
Низкая лабильность и высокая утомляемость синапсов. Синапсы могут проводить 50-100 нервных импульсов в секунду. Это в 5-10 раз меньше, чем максимальная частота ПД, которую могут воспроизводить нервные волокна при их электростимуляции. Если нервные волокна считаются практически неутомляемыми, то в синапсах утомление развивается весьма быстро. Это происходит из-за истощения запасов медиатора, энергетических ресурсов, развития стойкой деполяризации постсинаптической мембраны и т.д.
Высокая чувствительность синапсов к действию биологически активных веществ, лекарственных препаратов и ядов. Например, яд стрихнин блокирует функцию тормозных синапсов ЦНС, связываясь с рецепторами, чувствительными к медиатору глицину. Столбнячный токсин блокирует тормозные синапсы, нарушая выделение медиатора из пресинаптической терминали. В обоих случаях развиваются опасные для жизни организма явления. Примеры действия биологически активных веществ и ядов на передачу сигналов в нервно-мышечных синапсах рассмотрены выше.
Свойства облегчения и депрессии синоптической передачи. Облегчение синаптической передачи имеет место, когда нервные импульсы поступают к синапсу через короткое время (10-50 мс) друг за другом, т.е. достаточно часто. При этом в течение некоторого промежутка времени каждый последующий ПД, приходящий к пресинаптической мембране, вызывает увеличение содержания медиатора в синаптической щели, возрастание амплитуды ВПСП и увеличение эффективности синаптической передачи.
Одним из механизмов облегчения является накопление ионов Са 2 в пресинаптической терминали. Для удаления кальциевым насосом порции кальция, вошедшей в синаптическую терминаль при поступлении ПД, необходимо несколько десятков миллисекунд. Если в это время приходит новый потенциал действия, то новая порция кальция входит в терминаль и ее эффект на высвобождение нейромедиатора складывается с остаточным количеством кальция, которое кальциевый насос не успел удалить из нейроплазмы терминали.
Имеются и другие механизмы развития облегчения. Этот феномен в классических руководствах по физиологии называют также посттетанической потенциацией. Облегчение синаптической передачи имеет значение в функционировании механизмов памяти, для образования условных рефлексов и обучения. Облегчение передачи сигналов лежит в основе развития пластичности синапсов и улучшения их функций при частой активации.
Депрессия (угнетение) передачи сигналов в синапсах развивается при поступлении очень частых (для нервно-мышечного синапса более 100 Гц) нервных импульсов к пресинаптической мембране. В механизмах развития явления депрессии имеют значение истощение запасов медиатора в пресинаптической терминали, снижение чувствительности рецепторов постсинаптической мембраны к медиатору, развитие стойкой деполяризации постсинаптической мембраны, затрудняющих генерацию ПД на мембране постсинаптической клетки.
Электрические синапсы
Кроме синапсов с химической передачей возбуждения в организме есть синапсы с электрической передачей. Эти синапсы имеют очень узкую синаптическую щель и пониженное электрическое сопротивление между двумя мембранами. Благодаря наличию поперечных каналов между мембранами и низкому сопротивлению, электрический импульс легко проходит через мембраны. Электрические синапсы обычно характерны для однотипных клеток.
В результате воздействия раздражителя пресинаптический потенциал действия раздражает постсинаптическую мембрану, где возникает распространяющийся потенциал действия.
Характеризуются большей скоростью проведения возбуждения по сравнению с химическими синапсами и низкой чувствительностью к воздействию химических веществ.
Электрические синапсы бывают с одно- и двусторонней передачей возбуждения.
В организме встречаются и электрические тормозные синапсы. Тормозное влияние развивается за счет действия тока, который вызывает гиперполяризацию постсинаптической мембраны.
В смешанных синапсах может происходить передача возбуждения с помощью как электрических импульсов, так и медиаторов.
Синапс- специализированные структуры, которые обеспечивают передачу возбуждения с одной возбудимой клетки на другую. Понятие СИНАПС введено в физиологию Ч.Шеррингтоном (соединение, контакт). Синапс обеспечивает функциональную связь между отдельными клетками. Подразделяются на нервно-нервные, нервно-мышечные и синапсы нервных клеток с секреторными клетками (нервно-железистые). В нейроне выделяется три функциональных отдела: сома, дендрит, аксон. Поэтому между нейронами существуют все возможные комбинации контактов. Например, аксо-аксональный, аксо-соматический и аксо-дендритный.
Классификация.
1)по местоположению и принадлежности соответствующим структурам:
- периферические (нервно-мышечные, нейросекреторные, рецепторнонейрональные);
- центральные (аксо-соматические, аксо-дендритные, аксо-аксональные, сомато-дендритные. сомато-соматические);
2)механизму действия- возбуждающие и тормозящие;
3)способу передачи сигналов- химические, электрические, смешанные.
4)химические классифицируют по медиатору, с помощью которого осуществляется передача- холинергические, адренергические, серотонинергические, глицинергически. и т.д.
Строение синапса.
Синапс состоит из следующих основных элементов:
Пресинаптической мембраны (в нервно-мышечном синапсе - это концевая пластинка):
Постсинаптической мембраны;
Синаптической щели. Синаптическая щель заполнена олигосахаридсодержащей соединительной тканью, которая играет роль поддерживающей структуры для обеих контактирующих клеток.
Систему синтеза и освобождения медиатора.
Систему его инактивации.
В нервно-мышечном синапсе пресиниптическая мембрана-часть мембраны нервного окончания в области контакта его с мышечным волокном, постсинаптическая мембрана- часть мембраны мышечного волокна.
Строение нервно-мышечного синапса.
1 -миелинизированное нервное волокно;
2 -нервное окончание с пузырьками медиатора;
3 -субсинаптическая мембрана мышечного волокна;
4 -синаптическая щель;
5-постсинаптическая мембрана мышечного волокна;
6 -миофибриллы;
7 -саркоплазма;
8 -потенциал действия нервного волокна;
9 -потенциал концевой пластинки (ВПСП):
10 -потенциал действия мышечного волокна.
Часть постсинаптической мембраны, которая расположена напротив пресинаптической, называется субсинаптической мембраной. Особенностью субсинаптической мембраны является наличие в ней специальных рецепторов, чувствительных к определенному медиатору и наличие хемозависимых каналов. В постсинаптической мембране, за пределами субсинаптической, имеются потенциалозависимые каналы.
Механизм передачи возбуждения в химических возбуждающих синапсах . В 1936 году Дейл доказал, что при раздражении двигательного нерва в его окончаниях в скелетной мышце выделяется ацетилхолин. В синапсах с химической передачей возбуждение передается с помощью медиаторов (посредников) .Медиаторы – химическкие вещества, которые обеспечивают передачу возбуждения в синапсах. Медиатором в нервно-мышечном синапсе является ацетилхолин, в возбуждающих и тормозных нервно-нервных синапсах - ацетилхолин, катехоламины - адреналин, норадреналин, дофамин; серотонин; нейтральные аминокислоты - глутаминовая, аспарагиновая; кислые аминокислоты - глицин, гамма-аминомасляная кислота; полипептиды: вещество Р, энкефалин, соматостатин; другие вещества: АТФ, гистамин, простагландины.
Медиаторы в зависимости от их природы делятся на несколько групп:
Моноамины (ацетилхолин, дофамин, норадреналин,серотонин.);
Аминокислоты (гамма-аминомасляная кислота - ГАМК, глутаминовая кислота, глицин и др.);
Нейропептиды (вещество Р, эндорфины, нейротензин, АКТГ,ангиотензин, вазопрессин, соматостатин и др.) .
Накопление медиатора в пресинаптическом образовании происходит за счет его транспорта из околоядерной области нейрона с помощью быстрого акстока; синтеза медиатора, протекающего в синаптических терминалях из продуктов его расщепления; обратного захвата медиатора из синаптическои щели.
Пресинаптическое нервное окончание содержит структуры для синтеза нейромедиатора. После синтеза нейромедиатор упаковывается в везикулы. При возбуждении эти синаптические везикулы сливаются с пресинаптической мембраной и нейромедиатор высвобождается в синаптическую щель. Он диффундирует к постсинаптической мембране и связывается там со специфическим рецептором. В результате образования нейромедиатор-рецепторного комплекса постсинаптическая мембрана становится проницаемой для катионов и деполяризуется. Это приводит к возникновению возбуждающего постсинаптического потенциала и затем потенциала действия. Медиатор синтезируется в пресинаптической терминали из материала, поступающего сюда аксональным транспортом. Медиатор "инактивируется", т.е. либо расщепляется, либо удаляется из синаптической щели посредством механизма обратного транспорта в пресинаптическую терминаль.
Значение ионов кальция в секреции медиатора .
Секреция медиатора невозможна без участия в этом процессе ионов кальция. При деполяризации пресинаптической мембраны кальций входит в пресинаптическую терминаль через специфические потенциалозависимые кальциевые каналы в этой мембране. Концентрация кальция в аксоплазме 110 -7 М, при вхождении кальция и повышения его концентрации до 110 - 4 М происходит секреция медиатора. Концентрация кальция в аксоплазме после окончания возбуждения снижается работой систем: активного транспорта из терминали, поглощением митохондриями, связыванием внутриклеточными буферными системами. В состоянии покоя происходит нерегулярное опорожнение везикул, при этом происходит выход не только единичных молекул медиатора, но и выброс порций, квантов медиатора. Квант ацетилхолина включает примерно 10000 молекул.
Тоже результат активности маленьких нервных клеток. Но это необыкновенно нужная и сложная работа была бы невозможна без синапсов, которые обеспечивают взаимодействие нейронов и связывают их в единые нейронные сети.
Если перевести слово «синапс» с греческого, то получится «связь». Это и есть место связи, соединения двух нейронов. Казалось бы, что тут такого особенного в обычном соединении? Но именно синапсы делают возможным прохождение импульса по цепи нервных клеток и играют важную роль во всех .
Место синапсов в нервной системе
Одна из главных задач нейронов – сохранение и обработка, поступающей из внешнего мира информации. От органов чувств, мышц, связок и т. д. слабые электрические сигналы по нервным волокнам попадают в головной мозг, там они распространяются по нейронным цепям, создавая очаги возбуждения и связи между отдельными нейронами, центрами и отделами головного мозга. Так вкратце происходят все процессы в нашей психике: от простейших безусловных рефлексов, до сложнейшей мыслительной деятельности.
Распространение нервных импульсов происходит благодаря имеющимся у нейронов отросткам. Короткие и сильно разветвленные дендриты специализируются на приеме сигналов от других нейронов. У одной нервной клетки может быть до 1500 дендритов. А вот передающее нервное волокно – аксон – одно, но оно длинное и может достигать 1,5 метров. Соединяясь с отростками дендритов, аксон передает сигнал от одного нейрона к другому.
Но проблема в том, что напрямую импульс чаще всего пройти не может, так как между «ветвями» дендрита одной нервной клетки и аксоном другой есть щель – пространство, заполненное межклеточным веществом.
Происходит следующее: в процессе движения импульса в месте соединения нервных волокон возникает биохимическая реакция, образуется белковая молекула – нейротрансмиттер или медиатор (посредник) – и закупоривает щель, создавая своеобразный мостик для прохождения сигнала.
Так возникает то, что еще в 1897 году английский физиолог Ч. Шеррингтон назвал синапсом.
Структура синапса
Если учесть, что размер нервной клетки редко превышает 100 мкм, то место соединения передающего и принимающего волокон двух нейронов вообще микроскопическое. И тем не менее, синапс имеет сложное строение, включающее в себя три основных отдела:
- Нервное окончание «ветвей» дендрита, которое представляет собой микроскопическое утолщение, называемое пресинаптической мембраной. Это очень важная часть синапса, отвечающая за синтез белковых молекул.
- Аналогичное утолщение на отростках аксона. Оно имеет специальные рецепторы, позволяющие принимать сигналы от медиаторов. Это постсинаптическая мембрана.
- Синаптическая щель, в которой образуется медиатор – проводящая импульс белковая молекула. Эта часть синапса одновременно и препятствует прохождению сигнала, и является причиной возникновения молекул белков, которые не только играют роль «мостиков», но и участвуют в работе нервной системы и организма в целом.
Функции этих белковых соединений разнообразны, так как нейроны вырабатывают разные виды медиаторов, и их химический состав оказывает различное влияние на процессы в нервной системе. Причем влияние это настолько сильное, что оно во многом управляет психическими реакциями, а недостаток даже одного из белков может привести к серьезным заболеваниям, таким как болезнь Паркинсона или Альцгеймера.
Сейчас обнаружено и изучено более 60 видов нейротрансмиттеров с разными свойствами. Вот примеры некоторых из них:
- Норадреналин – гормон . Он обладает возбуждающим действием, повышает активность всех систем организма и добавляет чувство ярости в наше эмоциональное состояние.
- Серотонин. Его функции многообразны: от обеспечения процесса пищеварения до влияния на уровень сексуального влечения.
- Глутамат необходим для запоминания и сохранения информации, но его переизбыток токсичен и может вызвать гибель нервных клеток.
- Дофамин – гормон счастья, источник позитивных , дарующий состояние блаженства. И одновременно этот белок, как и многие другие, обеспечивает эффективность познавательных процессов. А его недостаток может вызвать состояние и привести к слабоумию.
Это далеко не все белки, которые вырабатывают нейроны, но даже такой пример позволяет оценить значение нейротрансмиттеров и роль синапсов в организации нормальной деятельности головного . Разрушение нервных связей в результате болезни или травмы может привести и к серьезным нарушениям психических функций.
Виды синапсов
Синапсы обеспечивают связи не только между нейронами головного мозга, но и с нервными клетками органов чувств, рецепторами, расположенными во внутренних органах, мышцах и связках. Поэтому существует большое разнообразие синапсов в зависимости от специализации нейронов, от характера их воздействия, от того белкового соединения, которое вырабатывается при прохождении импульса.
В нашей нервной системе существует два основных процесса, определяющих ее деятельность. Это возбуждение и торможение. В соответствии с ними и синапсы делятся на два вида:
- возбуждающие проводят сигналы, которые распространяют реакцию возбуждения нервных клеток;
- тормозящие обеспечивают прохождение нервного импульса, который передает нейронам «команду» торможения.
По месту расположения синапсы различаются:
- на центральные, расположенные в головном мозге;
- периферические, обеспечивающие связи нейронов за пределами мозга – в периферической нервной системе.
Передача импульсов через синаптическую щель тоже может проводиться разными способами, в соответствии с этим выделяют три вида синапсов:
- Химические синапсы расположены в коре головного мозга. Они проводят сигнал с помощью нейротрасмиттеров, которые образуются в результате биохимической реакции.
- Электрические – та часть синапсов, которые способны передавать электрический сигнал без посредников-медиаторов. Например, это касается нейронов, расположенных в зрительном рецепторе. В этом случае химическая реакция не происходит, и обмен сигналами осуществляется быстрее.
- Электрохимические синапсы сочетают в себе особенности обеих этих групп.
Также существует классификация синапсов по видам трансмиттеров. Например, если производится норадреналин, но синапсы эти называются адренергические, а если ацетилхолин, то – холинергические. Учитывая, что белков, вырабатываемых нейронами, несколько десятков видов, мы имеем очень объемную классификацию, которая здесь вряд ли уместна.
Синапсы и нейронные сети
Синапсы, устанавливая связи между проводящими нервными волокнами, обеспечивают возникновение и поддержание в рабочем состоянии нейронных цепей. Соединяясь и переплетаясь, они образуют сложные нейронные сети, по которым с огромной скоростью проносятся электрические импульсы.
По последним научным данным, только в коре головного мозга функционирует около 100 млрд нейронов. Каждый из них способен иметь до 10 000 синапсов, то есть связей с другими нервными клетками. И они могут обмениваться сигналами со скоростью 100 м/сек. Представляете, какой объем информации циркулирует в нашей нервной системе?
Результаты недавних исследований американских нейрофизиологов позволяют утверждать, что потенциальный объем памяти головного мозга человека измеряется петабайтами. 1 петабайт – 10 15 байт или 1 миллион гигабайт. И это сопоставимо с объемом информации, циркулирующей во всемирном интернет-пространстве. Поэтому когда не слишком радивый студент говорит, что у него распухла голова от полученных знаний и ничего больше туда впихнуть он не может, то стоит в этом усомниться.