Утомление нервно-мышечного препарата и утомление в целом организме
Под утомлением следует понимать временное понижение работоспособности клетки, органа или организма, которое возникает в результате работы и исчезает после отдыха.
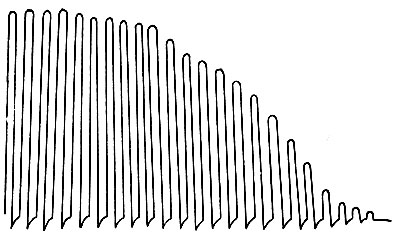
Утомление мышцы . Если на изолированную мышцу наносить одиночные ритмические раздражения индукционным током с частотой 1-2 раза в секунду и на барабане кимографа записывать ее сокращения (миограмма ), то можно отметить следующие явления. В первый период опыта наблюдается нарастание величины мышечных сокращений. Повышение работоспособности мышцы является результатом увеличения обменных процессов, возбудимости и лабильности. Затем в течение продолжительного времени наблюдается постоянная амплитуда сокращений мышцы. В дальнейшем отмечается постепенное снижение сократительного эффекта мышцы вплоть до отсутствия ее ответной реакции, что свидетельствует о развитии утомления (рис. 68).
Анализ миограмм показывает, что по мере развития утомления увеличивается продолжительность одиночного мышечного сокращения, главным образом за счет замедленного расслабления мышцы. В дальнейшем увеличивается латентный период сокращения и порог раздражения. При развитии утомления значительно нарастает хронаксия мышцы. Причинами возникающего в мышце утомления являются накопление продуктов обмена веществ (молочная, фосфорная кислоты и др.), уменьшение запаса кислорода и истощение энергетических ресурсов.
Утомление нервно-мышечного препарата . На нерв наносят достаточно сильные (или частые) раздражения и на барабане кимографа регистрируют кривую мышечных сокращений. При длительном раздражении нерва наблюдается постепенное снижение амплитуды сокращений и даже отсутствие ответной реакции мышцы (см. рис. 68). Ослабление силы наносимого раздражения или уменьшение его частоты также не сопровождается ответной реакцией мышцы, что свидетельствует о развитии утомления в нервно-мышечном препарате (см. рис. 68, Б).
Для того чтобы ответить на вопрос, в какой структуре нервно-мышечного препарата в первую очередь развивается утомление, перейдем к прямому раздражению мышцы стимулами исходной силы или частоты. В этом случае наблюдается восстановление механической реакции мышцы. Логично предположить, что утомление развилось либо в нерве, либо в мионевральном синапсе. Работами Н. Е. Введенского установлено, что нерв практически неутомляем. Следовательно, утомление в первую очередь развивается в области мионеврального синапса нервно-мышечного препарата лягушки, что связывают с истощением запасов медиатора в терминали нервного волокна. Кроме того, если сравнить лабильность различных образований нервно-мышечного препарата, то окажется, что функциональная подвижность мионеврального синапса самая низкая (рис. 69). В связи с этим в синапсе быстрее наступает утомление, как в структуре с более низкой лабильностью.
Отечественные физиологи И. М. Сеченов, И. П.Павлов, А. А. Ухтомский, Л. А. Орбели к проблеме утомления целостного организма подходили с позиций ведущей роли в ней центральной нервной системы. В организме, единство которого обеспечивается совместной деятельностью центральных и периферических нервных механизмов, утомление развивается раньше всего в нервных центрах.
На быстроту наступления утомления при длительном выполнении физической или умственной работы оказывают влияние режим жизни человека, условия его питания, сна, состояния центральной нервной системы, степень тренированности и т. д.
В конце прошлого столетия физиологи начали изучать отдельные проявления утомления. Итальянский ученый Моссо предложил эргографический метод исследования у человека процесса утомления, наступающего при мышечной работе. С помощью прибора эргографа было изучено влияние ритма выполняемой работы и величины поднимаемого груза на скорость возникновения утомления. Сущность эргографического метода состоит в том, что обследуемому предлагают путем разгибания и сгибания пальца верхней конечности, фиксированной в приборе, поднимать и опускать определенной величины груз в ритме ударов метронома. Движения пальца регистрируют на барабане кимографа. Кривую мышечных сокращений, записанную с помощью эргографа, называют эргограммой (рис. 70). Было установлено, что на развитие утомления в первую очередь оказывает влияние ритм выполняемой работы.
И. М. Сеченов изучал утомление, регистрируя сокращение мышц при поднятии груза на эргографе, сконструированном им самим. И. М. Сеченов обнаружил, что работоспособность утомленной руки во время ее отдыха восстанавливается полнее и лучше в том случае, если в этот период выполняет работу другая рука. Такое же влияние на работоспособность утомленной руки оказывает раздражение индукционным током афферентных нервов кисти другой руки, а также работа ногами, связанная с подъемом тяжести, и вообще двигательная активность.
Анализ установленных фактов позволил И. М. Сеченову прийти к заключению о том, что отдых, сопровождающийся умеренной работой мышечных групп, является более эффективным средством борьбы с утомлением двигательного аппарата, чем покой - пассивный отдых . В физиологии появилось понятие активный отдых .
Увеличение работоспособности после активного отдыха обусловлено повышением возбудимости нейронов центральной нервной системы под влиянием нервных импульсов, поступающих от проприорецепторов, а также адаптационно-трофическим воздействием симпатической нервной системы на утомленные мышечные группы (И. М. Сеченов, Л. А. Орбели).
Таким образом, лучшим способом борьбы с утомлением является смена формы труда, смена одного вида деятельности другим.
Утомлением называется временное понижение работоспособности клетки, органа или целого организма, наступающее в результате работы и исчезающее после отдыха.
Если длительно раздражать ритмическими электрическими стимулами изолированную мышцу, в которой подвешен небольшой груз, то амплитуда ее сокращений постепенно убывает, пока не дойдет до нуля. Полученная таким образом кривая называется кривой утомления мышцы . Измерив и суммировав высоту всех сокращений, можно узнать общую высоту подъёма груза, а умножив груз на эту величину, определить количество работы, выполненной мышцей до наступления полного утомления.
Наряду с изменением амплитуды сокращений при утомлении нарастает латентный период сокращения и увеличиваются пороги раздражения и хронаксия, т. е. понижается возбудимость. Следует, однако, подчеркнуть, что все эти изменения возникают не тотчас же после начала работы мышцы - существует некоторый период, в течение которого наблюдаются увеличение амплитуд сокращений и небольшое повышение возбудимости мышцы. При этом мышца становится легко растяжимой. В таких случаях говорят, что мышца «врабатывается», т. е. приспособляется к работе при заданном ритме и силе раздражения. При дальнейшем длительном раздражении наступает утомление мышечных волокон.
Понижение работоспособности изолированной из организма мышцы при ее длительном раздражении обусловлено двумя основными причинами. Первой из них является то, что во время сокращений в мышце накапливаются продукты обмена веществ (в частности, молочная кислота, образующаяся при расщеплении гликогена), оказывающие угнетающее влияние на работоспособность мышечных волокон. Часть этих продуктов, а также ионы калия диффундируют из волокон наружу в околоклеточное пространство и оказывают угнетающее влияние на способность возбудимой мембраны генерировать потенциалы действия.
Если изолированную мышцу, помещенную в раствор Рингера, довести длительным раздражением до полного утомления, то достаточно только сменить омывающую ее жидкость, чтобы восстановились сокращения мышцы.
Другой причиной развития утомления изолированной мышцы является постепенное истощение в ней энергетических запасов. При длительней работе изолированной мышцы происходит резкое уменьшение запасов гликогена, вследствие чего нарушаются процессы ресинтеза АТФ и креатинфосфата, необходимых для осуществления сокращения.
Рассматривая проблему утомления, следует подчеркнуть, что утомление изолированной скелетной мышцы при ее прямом раздражении является сугубо лабораторным феноменом и в естественных условиях существования организма утомление двигательного аппарата при длительной работе развивается совершенно отлично от того, что наблюдается в эксперименте. Обусловлено это не только тем, что в организме мышца непрерывно снабжается кровью и, следовательно, получает с кровью определённое количество питательных веществ (глюкозу, аминокислоты) и ocвобождается от продуктов обмена, нарушающих нормальную жизнедеятельность мышечных волокон. Главное отличие состоит в том, что в организме возбуждающие импульсы приходят к мышце с нерва. Нервно-мышечное coeдинение утомляется значительно раньше, чем мышечные волокна, в связи с чем блокирование передачи возбуждений с нерва на мышцу предохраняет последнюю от истощения, вызываемого длительной работой. В целостном организме еще раньше нервно-мышечных соединений утомляются при работе нервные центры.
Впервые И. М. Сеченов (1903) показал, что восстановление работоспособности утомленных мышц руки человека после длительной работы по подъему груза резко ускоряется, если в период отдыха производить работу другой рукой. Временное восстановление работоспособности мышц утомлённой руки может быть стигнутои при других видах двигательной активности, например при работе различных мышц нижних конечностей. В отличие от простого покоя такой отдых был назван И. М. Сеченовым активным. Сеченов рассматривал эти факты как доказательство того, что утомление прежде всего развивается в нервных центрах.
Убедительным доказательством роли изменения состояния нервных центров в развитии утомления в целом организме служат опыты с внушением. Так, исследуемый может длительно поднимать тяжелую гирю, если ему внушить, что в его руке находится легкая корзина. Напротив, если внушить исследуемому, поднимающему легкую корзину, что ему дана тяжелая гиря, то быстро развивается утомление. При этом изменения пульса, дыхания и газообмена находится в соответствии не с реальной, осуществляемой человеком работой, а с той, которая ему внушена (В. М. Василевский, Д. И. Шатенштейн).
Из изложенного выше следует, что утомление изолированной скелетной мышцы при ее прямом раздражении, утомление нервно-мышечного препарата при раздражении двигательного нерва и утомление двигательного аппарата в целом организме в условиях естественной деятельности сходны между собой лишь по своему внешнему выражению - уменьшению силы и величины мышечных сокращений.
По механизму же своего возникновения эти явления весьма значительно различаются.
Эргография . Для изучения мышечного утомления у человека в лабораторных условиях пользуются эргографами - приборами для записи амплитуды движения, ритмически выполняемого группой мышц.
Примером такого прибора может служить эргограф Моссо, записывающий движение нагруженного пальца при сгибании и разгибании и дающий суммарные сведения о работе собственного сгибателя этого пальца и общего сгибателя всех пальцев руки. Исследуемый, сгибая и разгибая палец, поднимает и опускает подвешенный к пальцу груз в ритме ударов метронома. Особый интерес представляют эргографы, воспроизводящие те или иные рабочие движения человека. Первым таким прибором был эргограф, примененный И. М. Сеченовым для изучения рабочих движений при пилке ручной пилой.
Меняя величину груза и частоту ударов метронома, можно установить тот ритм и груз, при которых данный индивидуум в данных условиях эксперимента выполняет наибольшую работу в кратчайший срок.
Утомление мышц
Основными показателями, характеризующими деятельность мышц, являются их сила и работоспособность.
Сила мышц. Сила - мера механического воздействия на мышцу со стороны других тел, которая выражается в ньютонах или кг-силах. При изотоническом сокращении в эксперименте сила определяется массой максимального груза, который мышца может поднять (динамическая сила), при изометрическом - максимальным напряжением, которое она может развить (статическая сила).
Работа мышц. При изометрическом и изотоническом сокращении мышца совершает работу.
Оценивая деятельность мышц, обычно учитывают только производимую ими внешнюю работу.
Работа мышцы, при которой происходит перемещение груза и костей в суставах называется динамической.
Утомление – это временное снижение работоспособности мышц в результате работы. Утомление изолированной мышцы можно вызвать ее ритмическим раздражением. В результате этого сила сокращений прогрессирующе уменьшается. Чем выше частота, сила раздражения и величина нагрузки, тем быстрее развивается утомление. При утомлении значительно изменяется кривая одиночного сокращения. Увеличивается продолжительность латентного периода, периода укорочения и особенно периода расслабления, но снижается амплитуда. Чем сильнее утомление мышцы, тем больше продолжительность этих периодов. В некоторых случаях полного расслабления не наступает. Развивается контрактура – это состояние длительного, непроизвольного сокращения мышцы.
Работа и утомление мышц исследуется с помощью эргографии. В прошлом веке, на основании опытов с изолированными мышцами, было предложено 3 теории мышечного утомления.
1. Теория Шиффа: утомление является следствием истощения энергетических запасов в мышце.
2. Теория Пфлюгера: утомление обусловлено накоплением в мышцах продуктов обмена.
3. Теория Ферворда: утомление объясняется недостатком кислорода в мышце.
Действительно, эти факторы способствуют утомлению в экспериментах на изолированных мышцах. В них нарушается ресинтез АТФ, накапливаются молочная и пировиноградная кислоты, недостаточно содержание кислорода. Однако в организме интенсивно работающие мышцы получают необходимый кислород, питательные вещества, освобождаются от метаболитов за счет усиления общего и регионального кровообращения. Поэтому были предложены другие теории утомления. В частности, определенная роль в утомлении принадлежит нервно-мышечным синапсам. Утомление в синапсе развивается из-за истощения запасов нейромидиатора. Однако главная роль в утомлении двигательного аппарата принадлежит моторным центрам центральной нервной системы. В прошлом веке И. М. Сеченов установил, что если наступает утомление мышц одной руки, то их работоспособность восстанавливается быстрее при работе другой рукой или ногами. Он считал, что это связано с переключением процессов возбуждения с одних двигательных центров на другие. Отдых с включением других мышечных групп н назвал активным. В настоящее время установлено, что двигательное утомление связано с торможением соответствующих нервных центров, в результате метаболических процессов в нейронах, ухудшением синтеза нейромидиатора, и угнетением синоптической передачи.
Основным морфо-функциональным элементом нервно-мышечного аппарата скелетных мышц является двигательная единица. Она включает мотонейрон спинного мозга с иннервируемыми его аксоном мышечными волокнами. Внутри мышцы этот аксон образует несколько концевых веточек. Каждая такая веточка образует контакт – нервно-мышечный синапс на отдельном мышечном волокне. Нервные импульсы, идущие от мотонейрона, вызывают сокращение определенной группы мышечных волокон. Двигательные единицы мелких мышц, осуществляющих тонкие движения (мышцы глаза, кисти), содержат небольшое количество мышечных волокон. В крупных их в сотни раз больше.
Все двигательные единицы в зависимости от функциональных особенностей делятся на 3 группы:
I. Медленные неутомляемые. Они образованы «красными» мышечными волокнами, в которых меньше миофибрилл. Скорость сокращения и сила этих волокон относительно небольшая, но они мало утомляемы. Поэтому их относят к тоническим. Регуляция сокращения таких волокон осуществляется небольшим количеством мотонейронов, аксоны которых имеют мало концевых веточек. Пример – камбаловидная мышца.
IIB. Быстро, легко утомляемые. Мышечные волокна содержат много миофибрилл и называются «белыми». Быстро сокращаются и развивают большую силу, но быстро утомляются. Поэтому их называют фазными. Мотонейроны этих двигательных единиц крупные, имеют толстый аксон с многочисленными концевыми веточками. Они генерируют нервные импульсы большой частоты. Например, мышцы глаза.
IIIА. Быстрые, устойчивые к утомлению. Занимают промежуточное положение.
Утомление - это временное снижение работоспособности мыши в результате работы. Утомление изолированной мышцы можно вызвать ее ритмическим раздражением. В результате этого сила сокращений прогрессирующе уменьшается (рис). Чем выше частота, сила раздражения, величина нагрузки тем быстрее развивается утомление. При утомлении значительно изменяется кривая одиночного сокращения. Увеличивается продолжительность латентного периода, периода укорочения и особенно периода расслабления, .но/снижается амплитуда (рис.) Чем сильнее утомление мышцы, тем больше продолжительность этих периодов. В некоторых случаях, полного расслабления не наступает, развивается контрактура. Это состояние непроизвольного длительного сокращения мышцы. Работа утомление мышц исследуются с помощью эргографии. В прошлом веке, на основании опытов с изолированными мышцами, было предложено 3 теории мышечного утомления.
1)Теория Шиффа: утомление является следствием истощения энергетических запасов, а мышце.
2. Теория Пфлюгера: утомление обусловлено накоплением в мышце продуктов обмена.
3. Теория Ферворна: утомление объясняется недостатком кислорода в мышце. Действительно эти факторы способствуют утомлению в экспериментах на изолированных мышцах. В них нарушается ресинтез ЛТФ. накапливается молочная и пировиноградная кислоты, недостаточно содержание кислорода. Однако в организме интенсивно работающие мышцы, получают необходимый кислород, питательные вещества, освобождаются от метаболитов за счет усиления общего и регионального кровообращения. Поэтому были предложены другие теории утомления. В частности, определенную роль в утомлении принадлежи? нервно-мышечным синапсам. Утомление в синапсе развивается из-за истощения запасов нейромедиатора. Однако главная роль, в утомлении двигательного аппарата принадлежит моторным центрам ЦНС. В прошлом веке И.М.Сеченов установил, что если наступает утомление мышц одной руки, то их работоспособность восстанавливается быстрее при работе другой рукой или ногами. Он считал, что это связано с переключением процессов возбуждения с одних двигательных центров на другие. Отдых с включением других мышечных групп он назвал активным. В настоящее время установлено, что двигательное утомление связано с торможением соответствующих нервных центров, в результате метаболических процессов в нейронах, ухудшением синтеза нейромедиаторов. и угнетением синаптической передачи.
Двигательные единицы
Основным морфо-функциональным элементов нервно-мышечного аппарата скелетных мышц является двигательная единица (ДЕ). Она включает мотонейрон спинного мозга с иннервируемым его аксоном мышечными волокнами. Внутри мышцы этот аксон образует несколько концевых веточек. Каждая такая веточка образует контакт - нервно-мышечный синапс на отдельном мышечном волокне. Нервные импульсы, идущие от мотонейрона, вызывают сокращения определенной группы мышечных волокон. Двигательные единицы мелких мышц, осуществляющих гонкие движения (мышцы глаза, кисти), содержат небольшое количество мышечных волокон. В крупных, их сотни раз больше. Все ДЕ в зависимости от функциональных особенностей делятся_на_3_группы:
I. Медленные неутомляемые. Они образованы красными мышечными волокнами, в которых меньше миофнбрил. Скорость сокращения и сила этих волокон относительно небольшие, но они мало утомляемы. Поэтому их относят тоническим. Регуляция сокращений таких, волокон осуществляется небольшим количеством мотонейронов, аксоны которых имеют мало концевых веточек. Пример, камбаловидная мышца. Н В. Быстрые, легко утомляемые. Мышечные волокна содержат много миофибрилл и называются "белыми". Быстро сокращаются и развивают большую силу, но быстро утомляются. Поэтому их называют фазными, Мотонейооны этих.."11^ самые крупные, имеют толстый аксон с многочисленными концевыми веточками. Они генерируют нервные импульсы большой частоты. Мышцы глаза. II А. Быстрые, устойчивые к утомлению. Занимают промежуточное положение.
Физиология гладких мышц
Гладкие мышцы имеются в стенках большинства органов пищеварения, сосудов, выводных протоков различных желёз мочевыводящей системы. Они являются непроизвольными и обеспечивают перистальтику органов пищеварения и мочевыводящей системы, поддержание тонуса сосудов. В отличие от скелетных, гладкие мышцы поперечности клетками чаще веретенообразной формы и небольших размеров, не имеющими поперечной почёрченности. Последнее связано с тем, что сократительный аппарат не обладает упорядоченным строением. Миофибриллы состоят из тонких нитей актина, которые идут в различных направлениях и прикрепляющихся к разным участкам сарколеммы. Миозиновые протофибриллы расположен рядом с актиновыми. Элементы саркоплазматического ретикулума не образуют систему трубочек. Отдельные мышечные клетки соединяются между собой контактами с низким электрическим сопротивлением - нексусами, что обеспечивает распространение возбуждения по всей гладкомышечной структуре. Возбудимость и проводимость гладких мышц ниже. чем скелетных. Мембранный потенциал составляет 40-60 мВ, так как мембрана ГМК имеет относительно высокую проницаемость для ионов натрия. Причем у многих гладких мышц МП не постоянен. Он периодически уменьшается и вновь возвращается к исходному уровню. Такие колебания называют медленными волнами (МВ). Когда вершина медленные полны достигает критического уровня деполяризации, на ней начинают генерироваться потенциалы действия. сопровождающиеся сокращениями (рис). МВ и ПД проводятся по гладким мышцам со скоростью всего от 5 до 50 см/сек. Такие гладкие мышцы называют спонтанно активными, т.е. они обладают автоматией. Например за счет такой активности происходит перистальтика кишечника. Водители ритма кишечной перистальтики расположены в начальных отделах соответствующих кишок.
Генерация ПД в ГМК обусловлена входом в них ионов кальция. Механизмы электромеханического сопряжения также отличаются. Сокращение развивается за счет кальция, входящего в клетку во время ПД, Опосредует связь кальция с укорочением миофибрилл важнейший клеточный белок - кальмодулин.
Кривая сокращения также отличается. Латентный период, период укорочения, а особенно расслабления значительно продолжительнее, чем у скелетных мышц. Сокращение длится несколько секунд. Гладким мышцам, в отличие от скелетных свойственно явление пластического тонуса. Это способность длительное время находится в состоянии сокращения без значительных энергозатрат и утомления. Благодаря этому свойству поддерживается форма внутренних органов и тонус сосудов. Кроме того, гладкомышечные клетки сами являются рецепторами растяжения. При их натяжении начинают генерироваться ПД, что приводит к сокращению ГМК. Это явление называется: миогенным механизмом регуляции сократительной активности. Изменение структуры мыши с возрастом
Анатомически у новорожденных имеются все скелетные мышцы, но относительно, веса тела они составляют всего 23% (у взрослого 44 %). Количество мышечных подокон в мышцах такое же как у взрослого. Однако микроструктура Мышечных волокон отличается.; Волокна меньше диаметром, в них больше ядер. По мере роста происходит. утолщение и удлинение волокон. Это происходит за счет утолщения миофибрилл, оттесняющих ядра на периферию. Размеры мышечных волокон стабилизируются к 20 годам.
Мышцы у детей эластичнее, чем у взрослых. Т.е. быстрее укорачиваются при сокращении и удлиняются при расслаблении. Возбудимость и лабильность мышц новорожденных, ниже чем взрослых, но с возрастом растет. У новорожденных даже во сне мышцы находятся в состоянии тонуса. Развитие различных групп мышц происходи г неравномерно. 84-5 лет более развиты мышцы предплечья, отстают в развитии мышцы кисти. Ускоренное согревание мышц кисти происходит в 6 - 7 лет. Причем разгибатели развиваются медленнее сгибателей. С возрастом изменяется соотношение тонуса мышц. В раннем детстве повышен тонус мышц кисти, разгибателей бедра т.д. постепенно распределение тонуса нормализуется.
Показатели силы и работы мыши в процессе роста
С возрастом сила мышечных сокращений увеличивается. Это объясняется не только увеличением мышечной массы, ни и совершенствованием двигательных рефлексов. Например, сила кисти с 5 до 16 лет возрастает в - 6 раз, мыши ног в 1 - 2,5 раза. Показатели силы до 10 лет больше у мальчиков. С 10 - 12 лет у девочек. Способность к быстрым и тонким движениям достигает оптимума к 14 годам, выносливость к 17. В 10 - 11 лет ребенок способен выполнять работу мощностью 100 вт, 18 -19- летние 250 - 300 вт.
Утомление - сложное явление, развивающееся во всем организме. Развивающееся в опыте утомление изолированной мышцы в связи с ее длительной работой выражается в постепенном уменьшении амплитуды сокращений, удлинении фазы расслабления, а также в том, что расслабление постепенно становится все менее полным - развивается контрактура. Специальные исследования обнаружили, что в утомленной мышце уменьшается возбудимость (порог раздражения повышается), удлиняется скрытый период (отрезок времени от момента начала раздражения мышцы до момента начала сокращения), увеличивается вязкость. Необходимо отметить, что эти признаки имеют место и при двигательной деятельности в мышцах всего организма.Нервно-мышечный препарат содержит в себе три элемента: мышечное волокно, нервно-мышечный синапс и нервное волокно. Опыт показывает, что при утомлении нервно-мышечного препарата изменение функциональных свойств наступает, в первую очередь, в нервно-мышечных синапсах, во вторую очередь, - непосредственно в мышечных волокнах. Что касается нервных проводников, то они, как впервые показал Н. Е. Введенский, практически «неутомимы». Изменение функциональных свойств нервно-мышечных синапсов выражается в нарушении процесса передачи возбуждения с нервных волокон на мышечные.Утомление организма как результат сдвигов в функциональном состоянии центральной нервной системы. Мышечная работа - это целостная деятельность всего организма. Функционирование организма как целого и его взаимодействие с внешним миром осуществляется посредством нервной системы при ведущей роли ее высшего отдела - коры больших полушарий.Нервная система наиболее чувствительна к изменениям внутренней среды. Такие факторы утомления, как накопление в крови продуктов работы клеток, уменьшение содержания в крови сахара, недостаток при некоторых условиях кислорода в крови, понижают работоспособность организма не прямо, а главным образом опосредствованно - через центральную нервную систему.Эти возможности коры больших полушарий и других отделов мозга, осуществляемые через посредство интрацентральных путей и вегетативных нервов, реализуются с помощью регулирующих влияний на все органы и ткани, в том числе также и на ЦНС. В активизации этих влияний ведущая роль принадлежит условнорефлекторным реакциям, возникающим при действии самых разнообразных сигнальных раздражителей.Утомление - процесс фазный, как и многие другие процессы в организме человека. В первой фазе возникает некоторое напряжение в деятельности физиологических систем. Устойчивое состояние может еще не нарушиться, но поддерживать его становится все труднее. Только современные математические приемы обработки результатов физиологических исследований с помощью компьютеров позволили «увидеть» эту фазу утомления.Во второй фазе уже отчетливо видны нарушения устойчивого состояния. При мышечной деятельности это проявляется в несогласованном снижении одних показателей и повышении других. Например, потребление кислорода может начать снижаться, а объемная скорость дыхания при этом возрастать. Это явный признак снижения эффективности и разбалансировки в деятельности вегетативных систем, характерный для утомления. При этом работа по-прежнему выполняется в том же объеме, с прежней интенсивностью: компенсаторные механизмы все еще справляются с удержанием необходимых функциональных свойств мышц. При умственной работе эта фаза обычно проявляется в увеличении числа ошибок, т.е. опять же в снижении эффективности, при сохранении скорости работы.Третья фаза - срыв устойчивого состояния. Разбалансировка в работе вегетативных систем быстро нарастает, их эффективность резко падает, и вслед за этим наступает отказ от работы («Не могу!»). Умственная работа, не требующая столь больших энергетических ресурсов, может при этом и продолжаться, однако ее неэффективность делает ее продолжение совершенно бессмысленным.Таким образом, чем дольше не наступает утомление при определенном уровне нагрузки либо чем выше уровень нагрузки, при котором наступает утомление, тем выше работоспособность человека.
14. Механизм проведения нервного импульса по безмиелиновым и миелиновым волокнам. Законы проведения возбуждения по нервам.
Механизмы проведения возбуждения по нервному волокнуМеханизм проведения возбуждения по нервным волокнам зависит от их типа. Существуют два типа нервных волокон: миелиновые и безмиелиновые.
Процессы метаболизма в безмиелиновых волокнах не обеспечивают быструю компенсацию расхода энергии. Распространение возбуждения будет идти с постепенным затуханием – с декрементом. Декрементное поведение возбуждения характерно для низкоорганизованной нервной системы. Возбуждение распространяется за счет малых круговых токов, которые возникают внутрь волокна или в окружающую его жидкость. Между возбужденными и невозбужденными участками возникает разность потенциалов, которая способствует возникновению круговых токов. Ток будет распространяться от «+» заряда к «-». В месте выхода кругового тока повышается проницаемость плазматической мембраны для ионов Na, в результате чего происходит деполяризация мембраны. Между вновь возбужденным участком и соседним невозбужденным вновь возникает разность потенциалов, что приводит к возникновению круговых токов. Возбуждение постепенно охватывает соседние участки осевого цилиндра и так распространяется до конца аксона.
В миелиновых волокнах благодаря совершенству метаболизма возбуждение проходит, не затухая, без декремента. За счет большого радиуса нервного волокна, обусловленного миелиновой оболочкой, электрический ток может входить и выходить из волокна только в области перехвата. При нанесения раздражения возникает деполяризация в области перехвата А, соседний перехват В в это время поляризован. Между перехватами возникает разность потенциалов, и появляются круговые токи. За счет круговых токов возбуждаются другие перехваты, при этом возбуждение распространяется сальтаторно, скачкообразно от одного перехвата к другому. Сальтаторный способ распространения возбуждения экономичен, и скорость распространения возбуждения гораздо выше (70-120 м/с), чем по безмиелиновым нервным волокнам (0,5–2 м/с).
Существует три закона проведения раздражения по нервному волокну.
Закон анатомо-физиологической целостности.
Проведение импульсов по нервному волокну возможно лишь в том случае, если не нарушена его целостность. При нарушении физиологических свойств нервного волокна путем охлаждения, применения различных наркотических средств, сдавливания, а также порезами и повреждениями анатомической целостности проведение нервного импульса по нему будет невозможно.
Закон изолированного проведения возбуждения.
Существует ряд особенностей распространения возбуждения в периферических, мякотных и безмякотных нервных волокнах.
В периферических нервных волокнах возбуждение передается только вдоль нервного волокна, но не передается на соседние, которые находятся в одном и том же нервном стволе.
В мякотных нервных волокнах роль изолятора выполняет миелиновая оболочка. За счет миелина увеличивается удельное сопротивление и происходит уменьшение электрической емкости оболочки.
В безмякотных нервных волокнах возбуждение передается изолированно. Это объясняется тем, что сопротивление жидкости, которая заполняет межклеточные щели, значительно ниже сопротивления мембраны нервных волокон. Поэтому ток, возникающий между деполяризованным участком и неполяризованным, проходит по межклеточным щелям и не заходит при этом в соседние нервные волокна.
Закон двустороннего проведения возбуждения.
Нервное волокно проводит нервные импульсы в двух направлениях – центростремительно и центробежно.
В живом организме возбуждение проводится только в одном направлении. Двусторонняя проводимость нервного волокна ограничена в организме местом возникновения импульса и клапанным свойством синапсов, которое заключается в возможности проведения возбуждения только в одном направлении.




