Каковы механизмы реализации генетической информации в клетке. Реализация генетической информации
После открытия принципа молекулярной организации такого вещества, как ДНК в 1953 году, начала развиваться молекулярная биология. Далее в процессе исследований ученые выяснили как рекомбенируется ДНК, ее состав и как устроен наш человеческий геном.
Каждый день на молекулярном уровне происходят сложнейшие процессы. Как устроена молекула ДНК, из чего она состоит? И какую роль играют в клетке молекулы ДНК? Расскажем подробно обо всех процессах, происходящих внутри двойной цепи.
Что такое наследственная информация?
Итак, с чего все начиналось? Еще в 1868 нашли в ядрах бактерий. А в 1928 г. Н. Кольцов выдвинул теорию о том, что именно в ДНК зашифрована вся генетическая информация о живом организме. Затем Дж. Уотсон и Ф. Крик нашли модель всем теперь известной спирали ДНК в 1953 году, за что заслужено получили признание и награду — Нобелевскую премию.
Что такое вообще ДНК? Это вещество состоит из 2 объединенных нитей, точнее спиралей. Участок такой цепочки с определенной информацией называется геном.

В ДНК хранится вся информация о том, что за белки будут формироваться и в каком порядке. Макромолекула ДНК — это материальный носитель невероятно объемной информации, которая записана строгой последовательностью отдельных кирпичиков — нуклеотидов. Всего нуклеотидов 4, они дополняют друг друга химически и геометрически. Этот принцип дополнения, или комплементарности, в науке будет описан позже. Это правило играет ключевую роль в кодировке и декодировании генетической информации.
Так как нить ДНК невероятно длинная, повторений в этой последовательности не бывает. У каждого живого существа собственная уникальная цепочка ДНК.
Функции ДНК
К функциям относятся хранение наследственной информации и ее
передача потомству. Без этой функции геном вида не мог бы сохраняться и развиваться на протяжении тысячелетий.
Организмы, которые претерпели серьезные
мутации генов, чаще не выживают или теряют способность производить потомство. Так происходит природная защита от вырождения вида.

Еще одна существенно важная функция — реализация хранимой информации. Клетка не может создать ни одного жизненно важного белка без тех инструкций, которые хранятся в двойной цепочке.
Состав нуклеиновых кислот
Сейчас уже достоверно известно, из чего состоят сами нуклеотиды — кирпичики ДНК. В их состав входят 3 вещества:
- Ортофосфорная кислота.
- Азотистое основание. Пиримидиновые основания — которые имеют только одно кольцо. К ним относят тимин и цитозин. Пуриновые основания, в составе которых присутствуют 2 кольца. Это гуанин и аденин.
- Сахароза. В составе ДНК — дезоксирибоза, В РНК — рибоза.
Число нуклеотидов всегда равно числу азотистых оснований. В специальных лабораториях расщепляют нуклеотид и выделяют из него азотистое основание. Так изучают отдельные свойства этих нуклеотидов и возможные мутации в них.
Уровни организации наследственной информации
Разделяют 3 уровня организации: генный, хромосомный и геномный. Вся информация, нужная для синтеза нового белка, содержится на небольшом участке цепочки — гене. То есть ген считается низший и самый простой уровень кодировки информации.

Гены, в свою очередь, собраны в хромосомы. Благодаря такой организации носителя наследственного материала группы признаков по определенным законам чередуются и передаются от одного поколения к другому. Надо заметить, генов в организме невероятно много, но информация не теряется, даже когда много раз рекомбенируется.
Разделяют несколько видов генов:
- по функциональному назначению выделяют 2 типа: структурные и регуляторные последовательности;
- по влиянию на процессы, протекающие в клетке, различают: супервитальные, летальные, условно летальные гены, а также гены мутаторы и антимутаторы.
Располагаются гены вдоль хромосомы в линейном порядке. В хромосомах информация сфокусирована не вразброс, существует определенный порядок. Существует даже карта, в которой отображены позиции, или локусы генов. Например, известно, что в хромосоме № 18 зашифрованы данные о цвете глаз ребенка .
А что же такое геном? Так называют всю совокупность нуклеотидных последовательностей в клетке организма. Геном характеризует целый вид, а не отдельную особь.
Каков генетический код человека?
Дело в том, что весь огромнейший потенциал человеческого развития заложен уже в период зачатия. Вся наследственная информация, которая необходима для развития зиготы и роста ребенка уже после рождения, зашифрована в генах. Участки ДНК и есть самые основные носители наследственной информации.
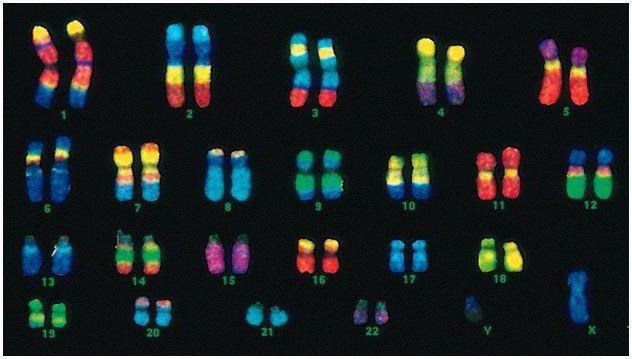
У человека 46 хромосом, или 22 соматические пары плюс по одной определяющей пол хромосоме от каждого родителя. Этот диплоидный набор хромосом кодирует весь физический облик человека, его умственные и физические способности и предрасположенность к заболеваниям. Соматические хромосомы внешне неразличимы, но несут они разную информацию, так как одна из них от отца, другая - от матери.
Мужской код отличается от женского последней парой хромосом — ХУ. Женский диплоидный набор — это последняя пара, ХХ. Мужчинам достается одна Х-хромосома от биологической матери, и затем она передается дочерям. Половая У-хромосома передается сыновьям.
Хромосомы человека значительно разнятся по размеру. Например, самая маленькая пара хромосом - №17. А самая большая пара - 1 и 3.
Диаметр двойной спирали у человека - всего 2 нм. ДНК настолько плотно закручена, что вмещается в маленьком ядре клетки, хотя ее длина будет достигать 2 метров, если ее раскрутить. Длина спирали — это сотни миллионов нуклеотидов.
Как передается генетический код?
Итак, какую роль играют в клетке молекулы ДНК при делении? Гены — носители наследственной информации - находятся внутри каждой клетки организма. Чтобы передать свой код дочернему организму, многие существа делят свое ДНК на 2 одинаковые спирали. Это называется репликацией. В процессе репликации ДНК расплетается и специальные «машины» дополняют каждую цепочку. После того как раздвоится генетическая спираль, начинает делиться ядро и все органеллы, а затем и вся клетка.
Но у человека другой процесс передачи генов - половой. Признаки отца и матери перемешиваются, в новом генетическом коде содержится информация от обоих родителей.
Хранение и передача наследственной информации возможны благодаря сложной организации спирали ДНК. Ведь как мы говорили, структура белков зашифрована именно в генах. Раз создавшись во время зачатия, этот код на протяжении всей жизни будет копировать сам себя. Кариотип (личный набор хромосом) не изменяется во время обновления клеток органов. Передача же информации осуществляется с помощью половых гамет — мужских и женских.
Передавать свою информацию потомству не способны только вирусы, содержащие одну цепочку РНК. Поэтому, чтобы воспроизводиться, им нужны клетки человека или животного.
Реализация наследственной информации
В ядре клетки постоянно происходят важные процессы. Вся информация, записанная в хромосомах, используется для построения белков из аминокислот. Но цепочка ДНК никогда не покидает ядро, поэтому здесь нужна помощь другого важного соединения = РНК. Как раз РНК способно проникнуть через мембрану ядра и взаимодействовать с цепочкой ДНК.
Посредством взаимодействия ДНК и 3 видов РНК происходит реализация всей закодированной информации. На каком уровне происходит реализация наследственной информации? Все взаимодействия происходят на уровне нуклеотидов. Информационная РНК копирует участок цепи ДНК и приносит эту копию в рибосому. Здесь начинается синтез из нуклеотидов новой молекулы.
Для того чтобы иРНК могла скопировать необходимую часть цепи, спираль разворачивается, а затем, по завершении процесса перекодировки, снова восстанавливается. Причем этот процесс может происходить одновременно на 2 сторонах 1 хромосомы.
Принцип комплементарности
Состоят из 4 нуклеотидов — это аденин (А), гуанин (G), цитозин (С), тимин (T). Соединены они водородными связями по правилу комплементарности. Работы Э. Чаргаффа помогли установить это правило, так как ученый заметил некоторые закономерности в поведении этих веществ. Э. Чаргафф открыл, что молярное отношение аденина к тимину равно единице. И точно так же отношение гуанина к цитозину всегда равно единице.
На основе его работ генетики сформировали правило взаимодействия нуклеотидов. Правило комплементарности гласит, что аденин соединяется только с тимином, а гуанин - с цитозином. Во время декодирования спирали и синтеза нового белка в рибосоме такое правило чередования помогает быстро найти необходимую аминокислоту, которая прикреплена к транспортной РНК.
РНК и его виды
Что такое наследственная информация? нуклеотидов в двойной цепи ДНК. А что такое РНК? В чем заключается ее работа? РНК, или рибонуклеиновая кислота, помогает извлекать информацию из ДНК, декодировать ее и на основе принципа комплементарности создавать необходимые клеткам белки.
Всего выделяют 3 вида РНК. Каждая из них выполняет строго свою функцию.
- Информационная (иРНК) , или еще ее называют матричная. Она заходит прямо в центр клетки, в ядро. Находит в одной из хромосом необходимый генетический материал для постройки белка и копирует одну из сторон двойной цепи. Копирование происходит снова по принципу комплементарности.
- Транспортная — это небольшая молекула, у которой на одной стороне декодеры-нуклеотиды, а на другой стороне соответствующие основному коду аминокислоты. Задача тРНК — доставить в «цех», то есть в рибосому, где синтезирует необходимую аминокислоту.
- рРНК — рибосомная. Она контролирует количество белка, который продуцируется. Состоит из 2 частей — аминокислотного и пептидного участка.
Единственное отличие при декодировании — у РНК нет тимина. Вместо тимина тут присутствует урацил. Но потом, в процессе синтеза белка, при ТРНК все равно правильно устанавливает все аминокислоты. Если же происходят какие-то сбои в декодировании информации, то возникает мутация.
Репарация поврежденной молекулы ДНК
Процесс восстановления поврежденной двойной цепочки называется репарацией. В процессе репарации поврежденные гены удаляются.

Затем необходимая последовательность элементов в точности воспроизводиться и врезается обратно в то же место на цепи, откуда было извлечено. Все это происходит благодаря специальным химическим веществам — ферментам.
Почему происходят мутации?
Почему некоторые гены начинают мутировать и перестают выполнять свою функцию — хранение жизненно необходимой наследственной информации? Это происходит из-за ошибки при декодировании. Например, если аденин случайно заменен на тимин.
Существуют также хромосомные и геномные мутации. Хромосомные мутации случаются, если участки наследственной информации выпадают, удваиваются либо вообще переносятся и встраиваются в другую хромосому.

Геномные мутации наиболее серьезны . Их причина - это изменение числа хромосом. То есть когда вместо пары — диплоидного набора присутствует в кариотипе триплоидный набор.
Наиболее известный пример триплоидной мутации — это синдром Дауна, при котором личный набор хромосом 47. У таких детей образуется 3 хромосомы на месте 21-й пары.
Известна также такая мутация, как полиплодия. Но полиплодия встречается только у растений.
Генетический код – способ записи в молекуле ДНК информации о количестве и порядке расположения аминокислот в белке.
Свойства:
Триплетность - одна аминокислота кодируется тремя нуклеотидами
Неперекрываемость - один и тот же нуклеотидне может входить одновременно в состав двух или более триплетов
Однозначность (специфичность) - определённый кодон соответствует только одной
Универсальность - генетический код работает одинаково в организмах разного уровня сложности - от вирусовдочеловека
Вырожденность (избыточность) - одной и той же аминокислоте может соответствовать несколько кодонов.
14.Этапы реализации наследственной информации у прокариот и эукариот.
Репликация (синтез) ДНК
Синтез ДНК всегда начинается в строго определенных точках. Фермент топоизомераза раскручивает спираль. Геликаза разрушает водородные связи между цепями ДНК и образует вилку репликаций. SSB-белки препятствуют повторному формированию водородных связей.
РНК-праймаза синтезирует короткие фрагменты РНК (праймеры),которые присоединяются к 3"-концу.
ДНК-полимераза начинают от праймера и синтезирует дочернюю цепь(5" 3")-
Направление синтеза одной цепи ДНК совпадает с направлением движения вилки репликаций, поэтому данная цепь синтезируется непрерывно. Здесь синтез идет быстро. Направление синтеза второй цепи противоположно напралению вилки репликаций. Поэтому синтез данной цепи происходит в виде отдельных участков и идет медленно (фрагменты Оказаки).
Созревание ДНК: отщепляется РНК-праймеры, достраиваются недостающие нуклеотиды, фрагменты ДНК соединяются с помощью лигазы. Топоизомераза раскручивает спираль.
Этапы реализации наследственной информации (у эукариот)
1.Транскрипция
2.Процессинг
3.Трансялция
4.Посттрансляционные изменения
Трансляция – синтез молекулы РНК на основе молекулы ДНК. Ключевой фермент – РНК-полимераза.
РНК-полимераза должна распознать промотер и взаимодействовать с ним. Промотер –особый участок ДНК, который располагается перед информативной частью гена. Взаимодействие с промотором необходимо для активации РНК-полимеразы. После активации РНК-полимераза обеспечивает разрыв водородных связей между цепями ДНК.
Синтез РНК всегда происходит по определенной кодогенной цепи ДНК.На этой цепи промотер располагается ближе к 3"-концу.
Синтез РНК происходит по принципам комплементарности и антипараллельности.
РНК-полимераза достигает стоп-кодона (терминатор или терминирующей кодон).Это является сигналом для прекращения синтеза. Фермент инактивируется, отделяется от ДНК при этом освобождается вновь синтезированная молекула ДНК – первичный трансткрипт – про-РНК. Восстанавливается исходная структура ДНК.
Особенности строения гена эукариот:
У эукариотов гены включают в себя различные по функции участки
А) Интроны- фрагменты ДНК (гена), которые не кодируют аминокислоты в белке
Б)Экзоны – участки ДНК, которые кодируют аминокислоты в белке.
Прирывистая природа гена была обнаружена Роберцом и Шарпом (Ноб. Премия 1903г).
Количество интронов и экзонов в разных генах сильно отличается.
Процессинг (созревание)
Происходит созревание первичного транскрипта и образуется зрелая молекула матричной РНК, которая может участвовать в синтезе белка на рибосомах.
На 5"- конце РНК формируется особый участок (структура) – КЭП или шапочка. КЭП обеспечивает взаимодействие с малой субъединицей рибосомы.
На 3"-конце РНК присоединяется от 100 до 200 молекул нуклеотидов, несущих аденин (полиА). При синтезе белка эти нуклеотиды постепенно отщепляется, разрушение полиА является сигналом для разрушения молекул РНК.
К некоторым нуклеотидам РНК присоединяется группа CH 3 – метилирование. Это увеличивает устойчивость ДНК к действию ферментов цитоплазмы.
Сплайсинг – происходит вырезание интронов и сшивание между собой экзонов. Фермент рестриктаза удаляет, лигаза- сшивает)
Зрелая матричная РНК включает в себя:
Лидер обеспечивает связывание матричной РНК с субъединицей рибосомы.
СК – стартовый кодон – одинаковый у всех матричных РНК, кодирует аминокислоту
Кодирующий участок – кодирует аминокислоты в белке.
Стоп-кодон – сигнал о прекращаемся синтезе белка.
Во время процессинга происходит жесткий отбор в цитоплазму из ядра выходит около 10% молекул от числа первичных транскриптов.
Альтернативный сплайсинг
У человека имеется 25-30 тысяч генов.
Однако у человека выделено около 100 тысяч белков.
Альтернативный сплайсинг – это ситуация, при которой в клетках разных тканей один и тот же ген обеспечивает синтез одинаковых молекул проРНК. В разных клетках по разному определяется количество и границы между экзонами и интронами. В результате из одинаковых первичных транскриптов получаются различные мРНК и синтезируются разные белки.
Альтернативный сплайсинг доказан примерно для 50% генов человека.
Трансляция – это процесс сборки пептидной цепи на рибосомах согласно информации, заложенной в иРНК.
1.Инициация (начало)
2.Элонгация (удлинение молекулы)
3.Терминация (конец)
Инициация.
Молекула матрРНК с помощью КЭПа контактирует с малой субъединицей рибосомы. С помощью лидера РНК связывается с субъединицей рибосомы. К стартовому кодону присоединяется транспРНК, которая несет транспортную кислоту метионин. Затем присоединяется большая субъединица рибосомы. В целой рибосоме формируется два активных центра: аминоацильный и пептидильный. Аминоакцильный свободен, а пептидильный занят тРНК с метионином.
Элонгация.
В аминоакцильный цент входит мРНК, антикодон которой соответствует кодируещему.
После этого рибосома сдвигается относительно мРНК на 1 кодон.При этом аминоакцильный центр освобождается. В пептидильном центре находится мРНК, соединяется с второй аминокислотой. Процесс циклически повторяется.
3.Терминация
В аминоацильный центр поступает стоп-кодон, который распознается специальным белком, это является сигналом для прекращения синтеза белка. Субъединицы рибосомы разъединяются, освобождая при этом мРНК и вновь синтезируется полипептид.
4.Пострансляционные изменения.
При трансляции образуется первичная структура полипептида.Этого недостаточно для выполнения функций белка, поэтому белок изменяется, что обеспечивает его активность.
Образуется:
А) вторичная структура (водородные связи)
Б)глобула – третичная структура (дисульфидные связи)
В) четвертичная структура – гемоглобин
Г)Гликозилирование – присоединение к белку остатков сахаров (антитела)
Д) расщепление большого полипептида на несколько фрагментов.
Различия в реализации наследственной информации прокариот и эукариот:
1.У прокариот отстутсвуют экзоны и интроны, поэтому отсутствуют этапы процессинга и сплайсинга.
2.У прокариот транскрипция и трансляция происходит одновременно, т.е. идет синтез РНК и уже начинается синтез ДНК.
3.У эукариот синтез различных видов РНК контролируется различными ферментами. У прокариот все типы РНК синтезируются одним ферментом
4.У эукариот каждый ген имеет свой собственный уникальный промотер, у прокариот один промотер может контролировать работу несколькихгенов.
5. Только у прокариот имеется система Оперона
Важнейшие функции организма - обмен веществ, рост, развитие, передача наследственности, движение и др. - осуществляются в результате множества химических реакций с участием белков, нуклеиновых кислот и других биологически активных веществ. При этом в клетках непрерывно синтезируются разнообразные соединения: строительные белки, белки-ферменты, гормоны. В ходе обмена эти вещества изнашиваются и разрушаются, а вместо них образуются новые. Поскольку белки создают материальную основу жизни и ускоряют все реакции обмена веществ, жизнедеятельность клетки и организма в целом определяется способностью клеток синтезировать специфические белки. Их первичная структура предопределена генетическим кодом в молекулеДНК.
Молекулы белков состоят из десятков и сотен аминокислот (точнее, из аминокислотных остатков). Например, в молекуле гемоглобина их около 600, и они распределены в четыре полипептидные цепочки; в молекуле рибонуклеазы таких аминокислот 124 и т. д.
Главная роль в определении первичной структуры белка принадлежит молекулам ДНК. Разные ее участки кодируют синтез разных белков, следовательно, одна молекула ДНК участвует в синтезе многих индивидуальных белков. Свойства белков зависят от последовательности аминокислот в полипептидной цепи. В свою очередь чередование аминокислот определяется последовательностью нуклеотидов в ДНК, и каждой аминокислоте соответствует определенный триплет. Экспериментально доказано, что, например, участок ДНК с триплетом ААЦ соответствует аминокислоте лейцину, триплет АЦЦ - триптофану, триплет АЦА-цистеину и т.д. Распределив молекулу ДНК на триплеты, можно представить, какие аминокислоты и в какой последовательности будут располагаться в молекуле белка. Совокупность триплетов составляет материальную основу генов, а каждый ген содержит информацию о структуре специфического белка (ген - это основная биологическая единица наследственности; в химическом отношении ген есть участок ДНК, включающий несколько сотен пар нуклеотидов).
Генетический код - исторически сложившаяся организация молекул ДНК и РНК, при которой последовательность нуклеотидов в них несет информацию о последовательности аминокислот в белковых молекулах. Свойства кода: триплетность (кодон), неперекрываемость (кодоны следуют друг за другом), специфичность (один кодон может определять в полииептидной цепи только одну аминокислоту), универсальность (у всех живых организмов один и тот же кодон обусловливает включение в полипептид одну и ту же аминокислоту), избыточность (для большинства аминокислот существует несколько кодонов). Триплеты, не несущие информации об аминокислотах, являются стоп триплетами, обозначающими место начала синтеза и-РНК. (В.Б. Захаров. Биология. Справочные материалы. М.,1997)
Поскольку ДНК находится в ядре клетки, а синтез белка происходит в цитоплазме, существует посредник, передающий информацию с ДНК на рибосомы. Таким посредником служит и РНК, на которую нуклеотидная последовательность переписывается, в точном соответствии с таковой на ДНК - по принципу комплементарности. Этот процесс получил название транскрипции и протекает как реакция матричного синтеза. Он характерен только для живых структур и лежит в основе важнейшего свойства живого - самовоспроизведения. Биосинтезу белка предшествует матричный синтез иРНК на нити ДНК. Возникшая при этом иРНК выходит из ядра клетки в цитоплазму, где на нее нанизываются рибосомы, сюда же с помощью тРЙК доставляются аминокислоты.
Синтез белка - сложный многоступенчатый процесс, в котором участвуют ДНК, иРНК, тРНК, рибосомы, АТФ и разнообразные ферменты. Вначале аминокисдоты в цитоплазме активируются с помощью ферментов и присоединяются к тРНК (к участку, где расположен нуклеотид ЦЦА). На следующем этапе идет соединение аминокислот в таком порядке, в каком чередование нуклеотидов с ДНК передано на иРНК. Этот этап называется трансляцией. На нити иРНК размещается не одна рибосома, а группа их - такой комплекс называется полисома (Н.Е. Ковалев, Л.Д. Шевчук, О.И. Щуренко. Биология для подготовительных отделений медицинских институтов).
Схема Биосинтез белка
Синтез белка состоит из двух этапов - транскрипции и трансляции.
I. Транскрипция (переписывание) - биосинтез молекул РНК, осуществляется в хромосомах на молекулах ДНК по принципу матричного синтеза. При помощи ферментов на соответствующих участках молекулы ДНК (генах) синтезируются все виды РНК (иРНК, рРНК, тРНК). Синтезируется 20 разновидностей тРНК, так как в биосинтезе белка принимают участие 20 аминокислот. Затем иРНК и тРНК выходят в цитоплазму, рРНК встраивается в субъединицы рибосом, которые также выходят в цитоплазму.
II. Трансляция (передача) - синтез полипептидных цепей белков, осуществляется в рибосомах. Она сопровождается следующими событиями:
1. Образование функционального центра рибосомы - ФЦР, состоящего из иРНК и двух субъединиц рибосом. В ФЦР всегда находятся два триплета (шесть нуклеотидов) иРНК, образующих два активных центра: А (аминокислотный) - центр узнавания аминокислоты и П (пептидный) - центр присоединения аминокислоты к пептидной цепочке.
2. Транспортировка аминокислот, присоединенных к тРНК, из цитоплазмы в ФЦР. В активном центре А осуществляется считывание антикодона тРНК с кодоном иРНК, в случае комплементарностн возникает связь, которая служит сигналом для продвижения (скачок) вдоль иРНК рибосомы на один триплет. В результате этого комплекс "кодон рРНК и тРНК с аминокислотой" перемещается в активный центр П, где и происходит присоединение аминокислоты к пептидной цепочке (белковой молекуле). После чего тРНК покидает рибосому.
3. Пептидная цепочка удлиняется до тех пор, пока не закончится трансляция и рибосома не соскочит с иРНК. На одной иРНК может умещаться одновременно несколько рибосом (полисома). Полипептидная цепочка погружается в канал эндоплазматиче-ской сети и там приобретает вторичную, третичную или четвертичную структуру. Скорость сборки одной молекулы белка, состоящего из 200-300 аминокислот, составляет 1-2 мин. Формула биосинтеза белка: ДНК (транскрипция) --> РНК (трансляция) --> белок.
Завершив один цикл, полисомы могут принять участие в синтезе новых молекул белка.
Отделившаяся от рибосомы молекула белка имеет вид нити, которая биологически неактивна. Биологически функциональной она становится после того, как молекула приобретает вторичную, третичную и четвертичную структуру, т. е. определенную пространственно специфическую конфигурацию. Вторичная и последующие структуры белковой молекулы предопределены в информации, заложенной в чередовании аминокислот, т. е. в первичной структуре белка. Иначе говоря, программа образования глобулы, ее уникальная конфигурация определяются первичной структурой молекулы, которая в свою очередь строится под контролем соответствующего гена.
Скорость синтеза белка обусловлена многими факторами: температурой среды, концентрацией водородных ионов, количеством конечного продукта синтеза, присутствием свободных аминокислот, ионов магния, состоянием рибосом и др.
1.5. Этапы реализации генетической информации в клетке
Принципиально важным свойством генетической информации является ее способность к переносу (передаче) как в пределах одной клетки, так и от родительской клетки к дочерним либо между клетками разных индивидуумов в процессах клеточного деления и размножения организмов (см. также гл. 3). Что касается направлений внутриклеточного переноса генетической информации, то в случае ДНК-содержащих организмов они связаны с процессами репликации молекул ДНК, т.е. с копированием информации (см. подразд. 1.2), либо с синтезом молекул РНК (транскрипцией) и образованием полипептидов (трансляцией) (рис. 1.14). Как известно, каждый из указанных процессов осуществляется на основе принципов матричности и комплементарности.
Сложившиеся представления о переносе генетической информации по схеме ДНК → РНК → белок принято называть "центральной догмой" молекулярной биологии. Наряду с этим (наиболее распространенным) направлением переноса, который иногда обозначают как "общий перенос", известна и другая форма реализации генетической информации ("специализированный перенос"), обнаруженная у РНК-содержащих вирусов. В этом случае наблюдается процесс, получивший название обратной транскрипции, при котором первичный генетический материал (вирусная РНК), проникший в клетку-хозяина, служит матрицей для синтеза комплементарной ДНК с помощью фермента обратной транскриптазы (ревертазы), кодируемой вирусным геномом. В дальнейшем возможна реализация информации синтезированной вирусной ДНК в обычном направлении. Следовательно,
специализированный перенос генетической информации осуществляется по схеме РНК → ДНК → РНК → белок.
Транскрипция является первым этапом общего переноса генетической информации и представляет собой процесс биосинтеза молекул РНК по программе ДНК. Принципиальный смысл этого процесса состоит в том, что информация структурного гена (либо нескольких расположенных рядом генов), записанная в форме нуклеотидной последовательности кодирующей нити ДНК в ориентации 3"→ 5", переписывается (транскрибируется) в нуклеотидную последовательность молекулы РНК, синтезируемой в направлении 5" → 3" на основе комплементарного соответствия дезоксирибонуклеотидов матричной нити ДНК рибонуклеотидам РНК (А-У, Г-Ц, Т-А, Ц-Г) (рис. 1.15). В качестве продуктов транскрипции (транскриптов) можно рассматривать все типы молекул РНК, участвующих в биосинтезе белков в клетке, — матричные (информационные) РНК (мРНК, или иРНК), рибосомные РНК (рРНК), транспортные РНК (тРНК), малые ядерные РНК (мяРНК).
Процесс транскрипции обеспечивается комплексным действием ряда ферментов, к числу которых относится РНК-полимераза, представляющая собой сложный белок, состоящий из нескольких субъединиц и способный выполнять несколько функций. В отличие от прокариот (бактерий), в клетках которых имеется РНК-полимераза лишь одного типа, обеспечивающая синтез разных молекул РНК, у эукариот установлено наличие ядерных РНК-полимераз трех типов (I, II, III), а также РНК-полимераз клеточных органелл, содержащих ДНК (митохондрий, пластид). РНК-полимераза I находится в ядрышке и участвует в синтезе большинства молекул рРНК, РНК-полимераза II обеспечивает синтез мРНК и мяРНК, а РНК-полимераза III осуществляет синтез тРНК и одного варианта молекул рРНК.
Транскрипция подразделяется на три основные стадии — инициацию (начало синтеза РНК), элонгацию (удлинение полинуклеотидной цепочки) и терминацию (окончание процесса).

Инициация транскрипции зависит от предварительного специфического связывания РНК-полимеразы с узнаваемой ею короткой нуклеотидной последовательностью в участке молекулы ДНК (промоторе), расположенном перед стартовой точкой структурного гена, с которой начинается синтез РНК. Промоторы разных структурных генов могут быть идентичными либо содержат отличающиеся друг от друга последовательности нуклеотидов, что, вероятно, определяет эффективность транскрибирования отдельных генов и возможности регуляции самого процесса транскрипции (см. также подразд. 1.6). Промоторы многих генов прокариот имеют в своем составе универсальную последовательность 5"-ТАТААТ-3" (блок Прибнова), которая располагается перед стартовой точкой на расстоянии порядка 10 нуклеотидов и распознается РНК-полимеразой. Другая относительно часто встречающаяся узнаваемая последовательность этих организмов (5"-ТТГАЦА-3") обычно обнаруживается на расстоянии примерно 35 нуклеотидов от стартовой точки. В геномах эукариот функцию узнавания для РНК-полимеразы II могут выполнять универсальные последовательности ТАТА (блок Хогнесса), ЦААТ и состоящие из повторяющихся нуклеотидов Г и Ц (ГЦ-мотивы). При этом та или иная промоторная область может содержать либо одну из указанных последовательностей либо комбинацию двух или трех таких последовательностей.
Специфическое прочное связывание РНК-полимеразы с тем или иным узнаваемым ею участком промоторной области позволяет ей начать процесс расплетания молекулы ДНК вплоть до стартовой точки, с которой она начинает осуществлять полимеризацию рибонуклеотидов с использованием в качестве матрицы однонитевого 3"-5"-фрагмента ДНК.
Дальнейшее расплетание ДНК структурного гена сопровождается удлинением синтезируемого полирибонуклеотида (элонгацией нити РНК), продолжающимся вплоть до достижения РНК-полимеразой области терминатора. Последний представляет собой нуклеотидную последовательность ДНК, которая узнается РНК-полимеразой при участии других белковых факторов терминации, что приводит к окончанию синтеза транскрипта и его отсоединению от матрицы. В большинстве случаев терминатор находится в конце структурного гена, обеспечивая синтез одной моногенной молекулы мРНК. При этом у прокариот возможен синтез полигенной молекулы мРНК, кодирующей синтез двух и большего числа полипептидных цепочек. Происходит непрерывное транскрибирование нескольких расположенных рядом друг с другом структурных генов, имеющих один общий терминатор. Полигенная мРНК может содержать в своем составе нетранслируемые межгенные области (спейсеры), разделяющие кодирующие участки для отдельных полипептидов, что, вероятно, обеспечивает последующее разделение и самих синтезируемых полипептидов.
Поскольку структурные гены эукариот имеют прерывистое (мозаичное) строение, то их транскрипция имеет специфические особенности, отличающие ее от транскрипции у прокариот. В случае эукариотического гена, кодирующего синтез полипептида, этот процесс начинается с транскрибирования всей нуклеотидной последовательности, содержащей как экзонные, так и интронные участки ДНК. Образовавшаяся при этом молекула мРНК, отражающая структуру всего мозаичного гена, которую называют гетерогенной ядерной РНК (гяРНК) либо проматричной РНК (про-мРНК), претерпевает затем процесс созревания (процессинг мРНК).
Процессинг состоит в ферментативном разрезании первичного транскрипта (гяРНК) с последующим удалением его интронных участков и воссоединением (сплайсингом) экзонных участков, формирующих непрерывную кодирующую последовательность зрелой мРНК, которая в дальнейшем участвует в трансляции генетической информации. В качестве примера можно рассмотреть схему процессинга мРНК, синтезируемой при транскрипции гена β-глобиновой цепочки (рис. 1.16), структура которого обсуждалась ранее (см. рис. 1.13).
В процессинге принимают участие и короткие молекулы мяРНК, состоящие примерно из 100 нуклеотидов, которые представляют собой последовательности, являющиеся комплементарными последовательностям на концах интронных участков гяРНК. Спаривание комплементарных нуклеотидов мяРНК и гяРНК способствует сворачиванию в петлю интронных участков и сближению соответствующих экзонных участков гяРНК, что, в свою очередь, делает их доступными разрезающему действию ферментов (нуклеаз). Следовательно, молекулы мяРНК обеспечивают правильность вырезания интронов из гяРНК.
Во время процессинга происходит также модификация 5"-и 3"-концов формирующейся зрелой молекулы мРНК. Принципиальный смысл этого процесса можно рассмотреть на схемах

процессинга гена β-глобина человека (см. рис. 1.16) и полной нуклеотидной последовательности зрелой мРНК, образующейся в результате этого процесса. Как видно из рис. 1.17, на 5"-конце последовательности имеется короткий нетранслируемый (лидирующий) участок, состоящий из 17 триплетов, которые маркированы цифрами со знаком "минус". Этот участок кодируется транскрибируемой (но нетранслируемой) областью первого экзона β-гена (заштрихована на рис. 1.16). Модификация этого участка состоит в образовании 5"-концевого кэпа (от англ, cap — колпачок, шапочка), представляющего собой остаток 7-метилгуанозина, присоединенный к соседнему нуклеотиду необычным способом (с помощью три-фосфатной связи). Предполагается, что основная функция кэпа связана с узнаванием специфической последовательности молекулы рРНК, входящей в состав рибосомы, что обеспечивает точное прикрепление всего лидирующего участка молекулы мРНК к определенному участку этой рибосомы и инициацию процесса трансляции. Возможно также, что кэп предохраняет зрелую мРНК от преждевременного ферментативного разрушения во время ее транспортировки из ядра в цитоплазму клетки.
Модификация 3 "-конца мРНК β-глобина, также имеющего короткую нетранслируемую последовательность, кодируемую соответствующей областью третьего экзона β-гена (см. рис. 1.16), связана с образованием полиаденилового (поли А) "хвоста" молекулы, состоящего из 100 — 200 последовательно соединенных остатков адениловой кислоты. Для действия фермента, осуществляющего полиаденилирование, не нужна матрица, но требуется присутствие на 3"-конце мРНК сигнальной последовательности ААУААА (см. рис. 1.17). Предполагается, что полиадениловый "хвост" обеспечивает транспорт зрелой мРНК к рибосоме, защищая ее от ферментативного разрушения, но сам постепенно разрушается ферментами цитоплазмы, отщепляющими один за другим концевые нуклеотиды.
Трансляция как очередной этап реализации генетической информации заключается в синтезе полипептида на рибосоме, при котором в качестве матрицы используется молекула мРНК (считывание информации в направлении 5" → 3"). Следует заметить, что в клетках прокариот, не имеющих настоящего ядра с оболочкой, хромосомный генетический материал (ДНК) практически находится в цитоплазме, что определяет непрерывный характер взаимосвязи процессов транскрипции и трансляции. Иными словами, образовавшийся лидирующий 5"-конец молекулы мРНК, синтез которой еще не завершен, уже способен вступать в контакт с рибосомой, инициируя синтез полипептида, т.е. транскрипция и трансляция идут одновременно. Что касается эукариот, то процессы транскрипции их ядерной генетической информации и ее трансляции должны быть разделены во времени в связи с процессингом молекул РНК и необходимостью их последующей упаковки и

Рис. 1.17. Нуклеотидная последовательность зрелой мРНК -глобинового гена человека. Последовательность начинается с 7-метилгуанозина на 5"-конце (кэп-сайт), за которым следует короткий нетранслируемый участок РНК. Первый транслируемый кодон (АУГ) выделен шрифтом и помечен цифрой 0, поскольку кодируемая им аминокислота (метионин) в дальнейшем выщепляется из полипептида (первой аминокислотой зрелого белка будет валин, кодируемый ГУГ). Выделены также стоп-кодон УАА (кодон 147), на котором заканчивается трансляция (полипептид состоит из 146 аминокислот), и сигнальная последовательность для полиаденилирования (ААУААА) на 3"-конце транспортировки из кариоплазмы в цитоплазму с участием специальных транспортных белков.
Как и в случае транскрипции, процесс трансляции можно условно подразделить на три основные стадии — инициацию, элонгацию и терминацию.
Для инициации трансляции принципиально важное значение имеет специфичность структурной организации группы идентичных рибосом (полирибосомы, или полисомы), которая может участвовать в синтезе первичной структуры определенной белковой молекулы (полипептида), кодируемой соответствующей мРНК. Как известно, отдельная рибосома представляет собой клеточную органеллу, состоящую из молекул рРНК, которые определяют ее специфичность, и из белков. В составе рибосомы имеются 2 структурные субъединицы (большая и малая), которые можно дифференцировать на основании их способности по-разному осаждаться при ультрацентрифугировании препаратов очищенных рибосом из разрушенных клеток, т. е. по коэффициенту седиментации (величине 5). При определенных условиях в клетке может происходить разделение (диссоциация) этих двух субъединиц либо их объединение (ассоциация).
Рибосомы прокариот, а также митохондрий и хлоропластов состоят из большой и малой субъединиц с величинами 505 и 305 соответственно, тогда как у эукариот эти субъединицы имеют другие размеры (605 и 405). Поскольку процесс трансляции более детально был исследован у бактерий, то чаще всего его рассматривают в связи со структурой рибосом этих организмов. Как видно из рис. 1.18, рибосома содержит 2 участка, имеющих прямое отношение к инициации трансляции, обозначенные как P-участок (аминоацильный) и Р- участок (пептидильный), специфичность которых определяется сочетанием соответствующих областей субъединиц 505 и 305. При диссоциации субъединиц рибосомы эти участки становятся "недостроенными", что приводит к изменению их функциональной специфичности.
В процессе трансляции участвуют также молекулы тРНК, функции которых состоят в транспортировке аминокислот из цитозоля (цитоплазматического раствора) к рибосомам. Молекула тРНК, имеющая вторичную структуру в форме "клеверного листа", содержит в своем составе тройку нуклеотидов (антикодон), которая обеспечивает ее комплементарное соединение с соответствующим кодоном (триплетом) молекулы мРНК, кодирующей синтез полипептида на рибосоме, и акцепторный участок (на 3"-конце молекулы), к которому присоединяется определенная аминокислота (см. рис. 1.7). Процесс присоединения каждой из 20 аминокислот к акцепторному концу соответствующей тРНК связан с ее активацией определенным вариантом фермента аминоацил-тРНК-


синтетазы с использованием энергии аденозинтрифосфатов (молекул АТФ). Образовавшийся при этом специфический комплекс тРНК и аминокислоты, который получил название аминоацил-тРНК, перемещается затем к рибосоме и участвует в синтезе полипептида.
Инициация трансляции обеспечивается точным соединением лидирующего 5"-конца молекулы мРНК с определенной областью малой субъединицы диссоциированной рибосомы таким образом, что в "недостроенном" Р-участке оказывается стартовый (инициирующий) кодон АУГ этой молекулы (рис. 1.19). Функциональная особенность такого Р-участка состоит в том, что он может быть занят только инициирующей аминоацил-тРНК с антикодоном УАЦ, которая у эукариот несет аминокислоту метионин, а у бактерий — формилметионин. Поскольку синтез пояипептида всегда начинается с N-конца и нарастает в направлении к С-концу, то все белковые молекулы, синтезируемые в клетках прокариот, должны начинаться с N-формилметионина, а у эукариот — с N-метионина. Однако, в дальнейшем эти аминокислоты ферментативно выщепляются во время процессинга белковой молекулы (см. рис. 1.17).
После образования инициирующего комплекса в "недостроенном" Р-участке (см. рис. 1.19) становится возможным воссоединение малой и большой субъединиц рибосомы, что приводит к "достраиванию" Р-участка и A-участка. Лишь после этого следующая аминоацил-тРНК может занимать A-участок на основе принципа
комплементарности ее антикодона соответствующему кодону мРНК, находящемуся в этом участке (см. рис. 1.19).
Процесс элонгации начинается с образования пептидной связи между инициирующей (первой в цепочке) и последующей (второй) аминокислотами. Затем происходит перемещение рибосомы на один триплет мРНК в направлении 5"→ 3", что сопровождается отсоединением инициирующей тРНК от матрицы (мРНК), от инициирующей аминокислоты и выходом ее в цитоплазму. При этом вторая по счету аминоацил-тРНК передвигается из A-участка в Р-участок, а освободившийся А -участок занимается следующей (третьей по счету) аминоацил-тРНК. Процесс последовательного передвижения рибосомы "триплетными шагами" по нити мРНК повторяется, сопровождаясь освобождением тРНК, поступающих в Р-участок, и наращиванием аминокислотной последовательности синтезируемого полипептида.
Терминация трансляции связана с вхождением одного из трех известных стоп-триплетов мРНК в Л-участок рибосомы. Поскольку такой триплет не несет информации о какой-либо аминокислоте, но узнается соответствующими белками терминации, то процесс синтеза полипептида прекращается и он отсоединяется от матрицы (мРНК).
После выхода из функционирующей рибосомы свободный 5"-конец мРНК может вступать в контакт со следующей рибосомой полисомной группы, инициируя синтез еще одного (идентичного) полипептида. Следовательно, рассмотренный рибосомный цикл последовательно повторяется с участием нескольких рибосом одной и той же полисомы, в результате чего синтезируется группа идентичных полипептидов.
Посттрансляционная модификация полипептида представляет собой завершающий этап реализации генетической информации в клетке, приводящий к превращению синтезированного полипептида в функционально активную молекулу белка. При этом первичный полипептид может претерпевать процессинг, состоящий в ферментативном удалении инициирующих аминокислот, отщеплении других (ненужных) аминокислотных остатков и в химической модификации отдельных аминокислот. Затем происходит процесс сворачивания линейной структуры полипептида за счет образования дополнительных связей между отдельными аминокислотами и формирование вторичной структуры белковой молекулы (рис. 1.20). На этой основе формируется еще более сложная третичная структура молекулы.
В случае белковых молекул, состоящих более чем из одного полипептида, происходит образование комплексной четвертичной структуры, в которой объединяются третичные структуры отдельных полипептидов. В качестве примера можно рассмотреть модель молекулы гемоглобина человека (рис. 1.21), состоящей из


двух α-цепочек и двух β-цепочек, которые формируют стабильную тетрамерную структуру с помощью водородных связей. Каждая из глобиновых цепочек содержит также молекулу тема, который в комплексе с железом способен связывать молекулы кислорода, обеспечивая их транспортировку эритроцитами крови.
Базисные термины и понятия: акцепторный конец тРНК; аминоацил-тРНК; антикодон; гяРНК (про-РНК); инициация транскрипции и трансляции; инициирующая аминоацил-тРНК и аминокислота; инициирующий кодон мРНК; комплементарность; кэп; лидирующий 5"-конец мРНК; матричность; модификация концов молекулы мРНК; моногенная молекула мРНК; мРНК (иРНК); мяРНК; обратная транскриптаза (ревертаза); обратная транскрипция; общий перенос; перенос (передача) информации; полигенная молекула мРНК; полипептид; полирибосома (полисома); посттрансляционная модификация полипептида; промотор; процессинг РНК и полипептида; рибосома; РНК-полимераза; рРНК; специализированный перенос; сплайсинг; стартовая точка транскрипции; терминатор; терминация транскрипции и трансляции; транскрипт; транскрипция генетической информации; трансляция генетической информации; тРНК; элонгация транскрипции и трансляции; A-участок рибосомы; Р-участок рибосомы.
Принципиально важным свойством генетической информации является ее способность к переносу (передаче) как в пределах одной клетки, так и от родительской клетки к дочерним либо между клетками разных индивидуумов в процессах клеточного деления и размножения организмов. Что касается направлений внутриклеточного переноса генетической информации, то в случае ДНК-содержащих организмов они связаны с процессами репликации молекул ДНК, т. е. с копированием информации, либо с синтезом молекул РНК (транскрипцией) и образованием полипептидов
(трансляцией) (рис. 5.15). Каждый из указанных процессов осуществляется на основе принципов матричности и комплементарности.
Рис. 5.15.
Сложившиеся представления о переносе генетической информации по схеме ДНК -? РНК -+ белок принято называть «центральной догмой» молекулярной биологии. Наряду с этим (наиболее распространенным) направлением переноса, который иногда обозначают как «общий перенос», известна и другая форма реализации генетической информации («специализированный перенос»), обнаруженная у РНК-содержащих вирусов. В этом случае наблюдается процесс, получивший название обратной транскрипции у при котором первичный генетический материал (вирусная РНК), проникший в клетку-хозяина, служит матрицей для синтеза комплементарной ДНК с помощью фермента обратной транскриптазы {ревертазы) у кодируемой вирусным геномом. В дальнейшем возможна реализация информации синтезированной вирусной ДНК в обычном направлении. Следовательно, специализированный перенос генетической информации осуществляется по схеме РНК->ДНК->РНК-?белок.
Транскрипция - первый этап общего переноса генетической информации, представляет собой процесс биосинтеза молекул РНК по программе ДНК. Принципиальный смысл этого процесса состоит в том, что информация структурного гена (либо нескольких расположенных рядом генов), записанная в форме нуклеотидной последовательности кодирующей нити ДНК в ориентации 3"->5 переписывается (транскрибируется) в нуклеотидную последовательность молекулы РНК, синтезируемой в направлении 5*-?З* на основе комплементарного соответствия дезоксирибонуклеотидов матричной нити ДНК риб о нуклеотидам РНК (A-У, Г-Ц, Т-А, Ц-Г) (рис. 5.16). В качестве продуктов транскрипции (транскриптов) можно рассматривать все типы молекул РНК, участвующих в биосинтезе белков в клетке: матричные (информационные) РНК (мРНК, или иРНК), рибосом- ные РНК (рРНК), транспортные РНК (тРНК), малые ядерные РНК (мяРНК).

Рис. 5.16.
Процесс транскрипции обеспечивается комплексным действием ряда ферментов, к числу которых относится РНК-полимераза, представляющая собой сложный белок, состоящий из нескольких субъединиц и способный выполнять несколько функций. В отличие от прокариот (бактерий), в клетках которых имеется РНК-полимераза лишь одного типа, обеспечивающая синтез разных молекул РНК, у эукариот установлено наличие ядерных РНК-полимераз трех типов (I, II, III), а также РНК-полимераз клеточных органелл, содержащих ДНК (митохондрий, пластид). РНК-полимераза I находится в ядрышке и участвует в синтезе большинства молекул рРНК, РНК-полимераза II обеспечивает синтез мРНК и мяРНК, а РНК-полимераза III осуществляет синтез тРНК и одного варианта молекул рРНК - 5SPHK.
Транскрипция подразделяется на три основные стадии: инициацию (начало синтеза РНК), элонгацию (удлинение полинуклеотидной цепочки) и терминацию (окончание процесса).
Инициация транскрипции зависит от предварительного специфического связывания РНК-полимеразы с узнаваемой ею короткой нуклеотидной последовательностью в участке молекулы ДНК (промоторе), расположенном перед стартовой точкой структурного гена, с которой начинается синтез РНК. Промоторы разных структурных генов могут быть идентичными либо содержат отличающиеся друг от друга последовательности нуклеотидов, что, вероятно, определяет эффективность транскрибирования отдельных генов и возможности регуляции самого процесса транскрипции. Промоторы многих генов прокариот имеют в своем составе универсальную последовательность 5-ТАТААТ-З" (блок Прибнова), которая располагается перед стартовой точкой на расстоянии порядка 10 нуклеотидов и распознается
РНК-полимеразой. Другая относительно часто встречающаяся узнаваемая последовательность прокариот (5-ТТГ.АЦА-3") обычно обнаруживается на расстоянии примерно 35 нуклеотидов от стартовой точки. В геномах эукариот функцию узнавания для РНК-полимеразы II могут выполнять универсальные последовательности ТАТА (блок Хогнесса), ЦААТ и состоящие из повторяющихся нуклеотидов Г и Ц (ГЦ-мотивы). При этом та или иная промоторная область может содержать либо одну из указанных последовательностей либо комбинацию двух или трех таких последовательностей.
Специфическое прочное связывание РНК-полимеразы с тем или иным узнаваемым ею участком промоторной области позволяет ей начать процесс расплетания молекулы ДНК вплоть до стартовой точки, с которой она начинает осуществлять полимеризацию рибонуклеотидов с использованием в качестве матрицы однонитевого 3"-5"-фрагмента ДНК.
Элонгация. Дальнейшее расплетание ДНК структурного гена сопровождается удлинением синтезируемого полирибонуклеотида (элонгацией нити РНК), продолжающимся вплоть до достижения РНК-полимеразой области терминатора.
Терминация - процесс прекращения репликации ДНК, происходящей посредством терминатора. Последний представляет собой нуклеотидную последовательность ДНК, которая узнается РНК-полимеразой при участии других белковых факторов терминации, что приводит к окончанию синтеза транскрипта и его отсоединению от матрицы. В большинстве случаев терминатор находится в конце структурного гена, обеспечивая синтез одной моногенной молекулы мРНК. При этом у прокариот возможен синтез полигенной молекулы мРНК, кодирующей синтез двух и более поли- пептидных цепочек. Происходит непрерывное транскрибирование нескольких расположенных рядом друг с другом структурных генов, имеющих один общий терминатор. Полигенная мРНК может содержать в своем составе нетранслируемые межгенные области (спейссры), разделяющие кодирующие участки для отдельных полипептидов, что, вероятно, обеспечивает последующее разделение и самих синтезируемых полипептидов.
Поскольку структурные гены эукариот имеют прерывистое (мозаичное) строение, их транскрипция имеет специфические особенности, отличающие ее от транскрипции у прокариот. В случае эукариотического гена, кодирующего синтез полипептида, этот процесс начинается с транскрибирования всей нуклеотидной последовательности, содержащей как экзон- ные, так и интронные участки ДНК. Образовавшаяся при этом молекула мРНК, отражающая структуру всего мозаичного гена, которую называют гетерогенной ядерной РНК (гяРНК) либо про-матричной РНК (про-мРНК), претерпевает затем процесс созревания (процессинг мРНК).
Процессинг состоит в ферментативном разрезании первичного транскрипта (гяРНК) с последующим удалением его интронных участков и воссоединением (сплайсингом) экзонных участков, формирующих непрерывную кодирующую последовательность зрелой мРНК, которая в дальнейшем участвует в трансляции генетической информации. В качестве примера можно рассмотреть схему процессинга мРНК, синтезируемой при транскрипции гена р-глобиновой цепочки (рис. 5.17).

Рис. 5.17.
В процессинге принимают участие и короткие молекулы мяРНК, состоящие примерно из 100 нуклеотидов, которые представляют собой последовательности, являющиеся комплементарными последовательностями на концах интронных участков гяРНК. Спаривание комплементарных нуклеотидов мяРНК и гяРНК способствует сворачиванию в петлю интронных участков и сближению соответствующих экзонных участков гяРНК, что, в свою очередь, делает их доступными разрезающему действию ферментов (нуклеаз). Следовательно, молекулы мяРНК обеспечивают правильность вырезания интронов из гяРНК.
Во время процессинга происходит также модификация 5 - и З"-концов формирующейся зрелой молекулы мРНК. Принципиальный смысл этого процесса можно рассмотреть на схемах процессинга гена р-глобина человека (рис. 5.17) и полной нуклеотидной последовательности зрелой мРНК, образующейся в результате этого процесса (рис. 5.18).

Рис. 5.18.
На 5’-конце последовательности (рис. 5.18) имеется короткий не- транслируемый (лидирующий) участок, состоящий из 17 триплетов, которые маркированы цифрами со знаком «минус». Этот участок кодируется транскрибируемой (но нетранслируемой) областью первого экзона р-гена (заштрихована на рис. 5.17). Модификация этого участка состоит в образовании 5"-концевого кэпа (от англ, cap - колпачок, шапочка), представляющего собой остаток 7-мстил|-уанозина, присоединенный к соседнему нуклеотиду необычным способом (с помощью трифосфатной связи). Предполагается, что основная функция кэпа связана с узнаванием специфической последовательности молекулы рРНК, входящей в состав рибосомы, что обеспечивает точное прикрепление всего лидирующего участка молекулы мРНК к определенному участку этой рибосомы и инициацию процесса трансляции. Возможно также, что кэп предохраняет зрелую мРНК от преждевременного ферментативного разрушения во время ее транспортировки из ядра в цитоплазму клетки.
Нуклеотидная последовательность зрелой мРНК р-глобинового гена человека начинается с 7-метилгуанозина на 5"-конце (кэп-сайт), за которым следует короткий нетранслируемый участок РНК. Первый транслируемый кодон (АУТ) выделен шрифтом и помечен цифрой 0, поскольку кодируемая им аминокислота (метионин) в дальнейшем выщепляется из полипептида (первой аминокислотой зрелого белка будет валин, кодируемый ГУГ). Выделены также стоп-кодон УАА (кодон 147), на котором заканчивается трансляция (полипептид состоит из 146 аминокислот), и сигнальная последовательность для полиаденилирования (ААУААА) на З’-конце.
Модификация З"-конца мРНК Р-глобина, также имеющего короткую нетранслируемую последовательность, кодируемую соответствующей областью третьего экзона p-гена (рис. 5.17), связана с образованием поли- аденилового (поли А) «хвоста» молекулы, состоящего из 100-200 последовательно соединенных остатков адениловой кислоты. Для действия фермента, осуществляющего полиаденилирование, нс нужна матрица, но требуется присутствие на З"-конце мРНК сигнальной последовательности ААУААА (рис. 5.18). Предполагается, что полиадениловый «хвост» обеспечивает транспорт зрелой мРНК к рибосоме, защищая ее от ферментативного разрушения, но сам постепенно разрушается ферментами цитоплазмы, отщепляющими один за другим концевые нуклеотиды.
Трансляция как очередной этап реализации генетической информации заключается в синтезе полипептида на рибосоме, при котором в качестве матрицы используется молекула мРНК (считывание информации в направлении 5 е -?З*)- Следует заметить, что в клетках прокариот, не имеющих настоящего ядра с оболочкой, хромосомный генетический материал (ДНК) находится в цитоплазме, что определяет непрерывный характер взаимосвязи процессов транскрипции и трансляции. Иными словами, образовавшийся лидирующий 5-конец молекулы мРНК, синтез которой еще не завершен, уже способен вступать в контакт с рибосомой, инициируя синтез полипептида, т. е. транскрипция и трансляция идут одновременно. Что касается эукариот, то процессы транскрипции их ядерной генетической информации и ее трансляции должны быть разделены во времени в связи с процессингом молекул РНК и необходимостью их последующей упаковки и транспортировки из кариоплазмы в цитоплазму с участием специальных транспортных белков.
Как и в случае транскрипции, процесс трансляции можно условно подразделить на три основные стадии: инициацию, элонгацию и терминацию.
Для инициации трансляции принципиально важное значение имеет специфичность структурной организации группы идентичных рибосом (полирибосомы, или полисомы), которая может участвовать в синтезе первичной структуры определенной белковой молекулы (полипептида), кодируемой соответствующей мРНК. Отдельная рибосома представляет собой клеточную органеллу, состоящую из молекул рРНК, которые определяют ее специфичность, и из белков. В составе рибосомы имеются две структурные субъединицы (большая и малая ), которые можно дифференцировать на основании их способности по-разному осаждаться при ультрацентрифугировании препаратов очищенных рибосом из разрушенных клеток, т. е. по коэффициенту седиментации (величине 5). При определенных условиях в клетке может происходить разделение (диссоциация) этих двух субъединиц либо их объединение (ассоциация).
Рибосомы прокариот, а также митохондрий и хлоропластов состоят из большой и малой субъединиц с величинами 505 и 305 соответственно, тогда как у эукариот эти субъединицы имеют другие размеры (605 и 405). Поскольку процесс трансляции более детально был исследован у бактерий, то чаще всего его рассматривают в связи со структурой рибосом этих организмов. Рибосома содержит два участка (рис. 5.19), имеющих прямое отношение к инициации трансляции, обозначенных как А -участок (аминоацильный) и P-участок (пептидильный), специфичность которых определяется сочетанием соответствующих областей субъединиц 505 и 305. При диссоциации субъединиц рибосомы эти участки становятся «недостроенными», что приводит к изменению их функциональной специфичности.

Рис. 5.19.
Р - пептидильный участок; А - аминоацильный участок
В процессе трансляции участвуют также молекулы тРНК, функции которых состоят в транспортировке аминокислот из цитозоля (цитоплазматического раствора) к рибосомам. Молекула тРНК, имеющая вторичную структуру в форме «клеверного листа», содержит тройку нуклеотидов (антикодон), которая обеспечивает ее комплементарное соединение с соответствующим кодоном (триплетом) молекулы мРНК, кодирующей синтез полипептида на рибосоме, и акцепторный участок (на З"-конце молекулы), к которому присоединяется определенная аминокислота (см. рис. 5.6). Процесс присоединения каждой из 20 аминокислот к акцепторному концу соответствующей тРНК связан с ее активацией определенным вариантом фермента аминоацил-тРНК-синтетазы с использованием энергии аденозинтрифос- фатов (молекул АТФ). Образовавшийся при этом специфический комплекс тРНК и аминокислоты, который получил название аминоацил-тРНК, перемещается затем к рибосоме и участвует в синтезе полипептида.
Инициация трансляции обеспечивается точным соединением лидирующего 5-конца молекулы мРНК с определенной областью малой субъединицы диссоциированной рибосомы таким образом, что в «недостроенном» P-участке оказывается стартовый (инициирующий) кодон АУТ этой молекулы (рис. 5.20, а). Функциональная особенность такого Р-участка состоит в том, что он может быть занят только инициирующей аминоацил- тРНК с антикодоном УАЦ, которая у эукариот несет аминокислоту метионин, у бактерий - формилметионин. Поскольку синтез полипептида всегда начинается с Анконца и нарастаег в направлении к С-концу, то все белковые молекулы, синтезируемые в клетках прокариот, должны начинаться с N-формилметионина, а у эукариот - с TV-метионина. Однако в дальнейшем эти аминокислоты ферментативно выщепляются во время процессинга белковой молекулы (см. рис. 5.18).

Рис. 5.20. Начальные этапы трансляции: а - инициирующий комплекс; б, в- элонгация
После образования инициирующего комплекса в «недостроенном» Л-участке (рис. 5.20) становится возможным воссоединение малой и большой субъединиц рибосомы, что приводит к «достраиванию» Р -участка и Л-участка. Лишь после этого следующая аминоацил-тРНК может занимать /4-участок на основе принципа комплементарности ее антикодона соответствующему кодону мРНК, находящемуся в этом участке (рис. 5.20, б, в).
Процесс элонгации начинается с образования пептидной связи между инициирующей (первой в цепочке) и последующей (второй) аминокислотами. Затем происходит перемещение рибосомы на один триплет мРНК в направлении 5"-?З", сопровождаемое отсоединением инициирующей тРНК от матрицы (мРНК), от инициирующей аминокислоты и выходом ее в цитоплазму. При этом вторая по счету аминоацил-тРНК передвигается из Л-участка в Р-участок, а освободившийся /4-участок занимается следующей (третьей по счету) аминоацил-тРНК. Процесс последовательного передвижения рибосомы «триплетными шагами» по нити мРНК повторяется, сопровождаясь освобождением тРНК, поступающих в P-участок, и наращиванием аминокислотной последовательности синтезируемого полипептида.
Терминация трансляции связана с вхождением одного из трех известных стоп-триплетов мРНК в /4-участок рибосомы. Поскольку такой триплет не несет информации о какой-либо аминокислоте, но узнается соответствующими белками терминации, то процесс синтеза полипептида прекращается и он отсоединяется от матрицы (мРНК).
После выхода из функционирующей рибосомы свободный 5-конец мРНК может вступать в контакт со следующей рибосомой полисомной группы, инициируя синтез еще одного (идентичного) полипептида. Следовательно, рассмотренный рибосомный цикл последовательно повторяется с участием нескольких рибосом одной и той же полисомы, в результате чего синтезируется группа идентичных полипептидов.
Посттранашционная модификация полипептида представляет собой завершающий этап реализации генетической информации в клетке, приводящий к превращению синтезированного полипептида в функционально активную молекулу белка. При этом первичный полипептид может претерпевать процессинг, состоящий в ферментативном удалении инициирующих аминокислот, отщеплении других (ненужных) аминокислотных остатков и в химической модификации отдельных аминокислот. Затем происходят процесс сворачивания линейной структуры полипептида за счет образования дополнительных связей между отдельными аминокислотами и формирование вторичной структуры белковой молекулы (рис. 5.21). На этой основе формируется еще более сложная третичная структура молекулы.

Рис. 5.21.

Рис. 5.22.
В случае белковых молекул, состоящих более чем из одного полипептида, происходит образование комплексной четвертичной структуры, в которой объединяются третичные структуры отдельных полипептидов. В качестве примера можно рассмотреть модель молекулы гемоглобина человека (рис. 5.22), состоящей из двух а-цепочск и двух (J-цепочек, которые формируют стабильную тетрамерную структуру с помощью водородных связей.
Каждая из глобиновых цепочек содержит также молекулу гема, который в комплексе с железом способен связывать молекулы кислорода, обеспечивая их транспортировку эритроцитами крови.




